29 Population Genetics: How population evolve
By the end of this section, you will be able to:
- Define population genetics and describe how scientists use population genetics in studying population evolution
- Describe the different types of variation in a population
- Describe the different forces of evolution
- Explain how each evolutionary force can influence a population’s allele frequencies
People did not understand the mechanisms of inheritance, or genetics, at the time Charles Darwin and Alfred Russel Wallace were developing their idea of natural selection. This lack of knowledge was a stumbling block to understanding many aspects of evolution. The predominant (and incorrect) genetic theory of the time, blending inheritance, made it difficult to understand how natural selection might operate. Darwin and Wallace were unaware of the Austrian monk Gregor Mendel’s 1866 publication “Experiments in Plant Hybridization”, which came out not long after Darwin’s book, On the Origin of Species. Scholars rediscovered Mendel’s work in the early twentieth century at which time geneticists were rapidly coming to an understanding of the basics of inheritance. Initially, the newly discovered particulate nature of genes made it difficult for biologists to understand how gradual evolution could occur. However, over the next few decades scientists integrated genetics and evolution in what became known as the modern synthesis—the coherent understanding of the relationship between natural selection and genetics that took shape by the 1940s. This concept is generally accepted today. In short, the modern synthesis describes how evolutionary processes, such as natural selection, can affect a population’s genetic makeup, and, in turn, how this can result in the gradual evolution of populations and species. The theory also connects population change over time (microevolution), with the macroevolution processes that gave rise to new species, novel adaptations, and higher-order patterns of diversity.
Variation in populations
Populations often exhibit a wide range of phenotypic variation, which refers to the observable differences in physical and physiological traits among individuals. These variations can include differences in size, coloration, behavior, and other measurable characteristics. Phenotypic variation arises from the interaction between an individual’s genetic makeup and environmental factors, influencing how certain traits are expressed.
Understanding the sources of phenotypic variation within a population is crucial for predicting how populations will respond to different evolutionary pressures. For example, certain phenotypes may provide a survival or reproductive advantage in specific environments, leading to an increase in the frequency of those traits over time through natural selection.
Phenotypic variation can occur both within populations and between populations of the same species. Within-population variation refers to the differences observed among individuals in the same population. This variation provides the raw material for evolution, as it allows natural selection to favor advantageous traits. Between-population variation occurs when different populations of the same species exhibit distinct phenotypic traits. This can result from geographical separation, environmental differences, or historical factors that lead to divergent evolutionary paths. Studying both types of variation helps scientists understand how species adapt to different environments and how new species may eventually arise.
Underlying the phenotypic variation is genetic variation, which involves differences in the alleles among individuals in a population. Genetic variation is introduced through mutations, gene flow, and sexual reproduction, leading to new combinations of genes. When a population has multiple forms or alleles of a particular gene, it is said to be polymorphic.
This is where population genetics comes into play. Population genetics is the branch of biology that focuses on the study of allele frequencies within populations and how they change over time under the influence of evolutionary forces such as natural selection, genetic drift, mutation, and gene flow. Allele frequencies refer to how common a particular allele is in the population’s gene pool compared to other alleles of the same gene.
By examining allele frequencies, population genetics provides insights into the genetic structure of populations and how genetic variation contributes to evolutionary processes. For example, if a specific allele confers a survival advantage, its frequency may increase in the population over successive generations due to natural selection.
It’s important to note that not all traits are heritable. Acquired traits, such as muscle strength gained from regular exercise, are generally not passed on to offspring because they do not alter an organism’s genetics. Natural selection cannot act on these traits since they are not encoded in the DNA.
Population genetics bridges the gap between genetic variation and evolutionary change by analyzing how allele frequencies shift within and between populations. Changes in allele frequencies can lead to significant evolutionary outcomes, such as adaptation to new environments or the emergence of new species.
Population Genetics
Recall that a gene for a particular character may have several alleles, or variants, that code for different traits associated with that character. For example, in the ABO blood type system in humans, three alleles determine the particular blood-type carbohydrate on the surface of red blood cells. Each individual in a population of diploid organisms can only carry two alleles for a particular gene, but more than two may be present in the individuals that comprise the population. Mendel followed alleles as they were inherited from parent to offspring. In the early twentieth century, biologists in the area of population genetics began to study how selective forces change a population through changes in allele and genotypic frequencies.
The allele frequency (or gene frequency) is the rate at which a specific allele appears within a population. Until now we have discussed evolution as a change in the characteristics of a population of organisms, but behind that phenotypic change is genetic change. In population genetics, scientists define the term evolution as a change in the allele’s frequency in a population. Using the ABO blood type system as an example, the frequency of one of the alleles, IA, is the number of copies of that allele divided by all the copies of the ABO gene in the population. For example, a study in Jordan1 found a frequency of IA to be 26.1 percent. The IB and I0 alleles comprise 13.4 percent and 60.5 percent of the alleles respectively, and all of the frequencies added up to 100 percent. A change in this frequency over time would constitute evolution in the population.
The allele frequency within a given population can change depending on environmental factors; therefore, certain alleles become more widespread than others during the natural selection process. Natural selection can alter the population’s genetic makeup. An example is if a given allele confers a phenotype that allows an individual to better survive or have more offspring. Because many of those offspring will also carry the beneficial allele, and often the corresponding phenotype, they will have more offspring of their own that also carry the allele, thus, perpetuating the cycle. Over time, the allele will spread throughout the population. Some alleles will quickly become fixed in this way, meaning that every individual of the population will carry the allele, while detrimental mutations may be swiftly eliminated if derived from a dominant allele from the gene pool. The gene pool is the sum of all the alleles in a population.
Sometimes, allele frequencies within a population change randomly with no advantage to the population over existing allele frequencies. We call this phenomenon genetic drift. Natural selection and genetic drift usually occur simultaneously in populations and are not isolated events. It is hard to determine which process dominates because it is often nearly impossible to determine the cause of change in allele frequencies at each occurrence. We call an event that initiates an allele frequency change in an isolated part of the population, which is not typical of the original population, the founder effect. Natural selection, random drift, and founder effects can lead to significant changes in a population’s genome.
Lecture Video: Population genetics
Forces of Evolution
To under stand how evolution occurs, it’s essential to understand the forces of evolution that act upon genetic variation within a population. There are five primary forces of evolution:
- Genetic Drift
- Gene Flow (Migration)
- Mutation
- Gene Flow (Migration)
- Non-Random Mating
These evolutionary forces are not mutually exclusive, but can shape the genetic structure of populations in different ways. Each one is discussed in turn below.
Genetic Drift
Lecture Video: Genetic drift, founder events, and genetic bottlenecks.
The theory of natural selection stems from the observation that some individuals in a population are more likely to survive longer and have more offspring than others; thus, they will pass on more of their genes to the next generation. A big, powerful male gorilla, for example, is much more likely than a smaller, weaker one to become the population’s silverback, the pack’s leader who mates far more than the other males of the group. The pack leader will father more offspring, who share half of his genes, and are likely to also grow bigger and stronger like their father. Over time, the genes for bigger size will increase in frequency in the population, and the population will, as a result, grow larger on average. That is, this would occur if this particular selection pressure, or driving selective force, were the only one acting on the population. In other examples, better camouflage or a stronger resistance to drought might pose a selection pressure.
Another way a population’s allele and genotype frequencies can change is genetic drift (Figure 1), which is simply the effect of chance. By chance, some individuals will have more offspring than others—not due to an advantage conferred by some genetically-encoded trait, but just because one male happened to be in the right place at the right time (when the receptive female walked by) or because the other one happened to be in the wrong place at the wrong time (when a fox was hunting).

Small populations are more susceptible to the forces of genetic drift. Large populations, alternatively, are buffered against the effects of chance. If one individual of a population of 10 individuals happens to die at a young age before it leaves any offspring to the next generation, all of its genes—1/10 of the population’s gene pool—will be suddenly lost. In a population of 100, that’s only 1 percent of the overall gene pool; therefore, it is much less impactful on the population’s genetic structure.
LINK TO LEARNING
Watch an animation of random sampling and genetic drift in action.
Natural events, such as an earthquake disaster that kills—at random—a large portion of the population, can magnify genetic drift. Known as the bottleneck effect, it results in suddenly wiping out a large portion of the genome (Figure 2). At once, the survivors’ genetic structure becomes the entire population’s genetic structure, which may be very different from the pre-disaster population.
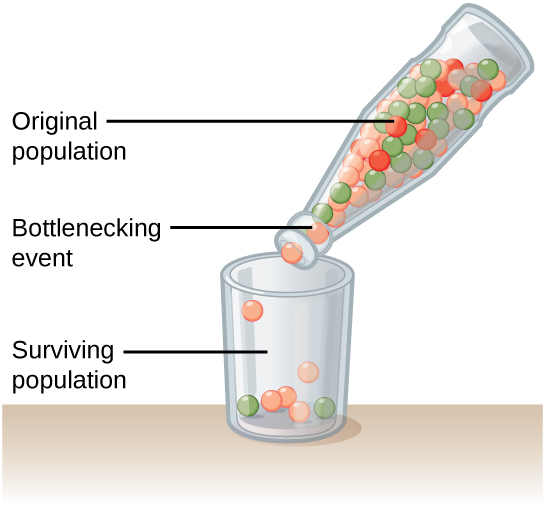
Another scenario in which populations might experience a strong influence of genetic drift is if some portion of the population leaves to start a new population in a new location or if a physical barrier divides a population. In this situation, those individuals are an unlikely representation of the entire population, which results in the founder effect. The founder effect occurs when the genetic structure changes to match that of the new population’s founding fathers and mothers. Researchers believe that the founder effect was a key factor in the genetic history of the Afrikaner population of Dutch settlers in South Africa, as evidenced by mutations that are common in Afrikaners but rare in most other populations. This is probably because a higher-than-normal proportion of the founding colonists carried these mutations. As a result, the population expresses unusually high incidences of Huntington’s disease (HD) and Fanconi anemia (FA), a genetic disorder known to cause blood marrow and congenital abnormalities—even cancer.1
VISUAL CONNECTION
SCIENTIFIC METHOD CONNECTION
Testing for the bottleneck effect
Question: How do natural disasters affect a population’s genetic structure?
Background: When an earthquake or hurricane suddenly wipes out much of a population, the surviving individuals are usually a random sampling of the original group. As a result, the population’s genetic makeup can change dramatically. We call this phenomenon the bottleneck effect.
Hypothesis: Repeated natural disasters will yield different population genetic structures; therefore, each time one runs this experiment the results will vary.
Test the hypothesis: Count out the original population using different colored beads. For example, red, blue, and yellow beads might represent red, blue, and yellow individuals. After recording the number of each individual in the original population, place them all in a bottle with a narrow neck that will only allow a few beads out at a time. Then, pour 1/3 of the bottle’s contents into a bowl. This represents the surviving individuals after a natural disaster kills a majority of the population. Count the number of the different colored beads in the bowl, and record it. Then, place all of the beads back in the bottle and repeat the experiment four more times.
Analyze the data: Compare the five populations that resulted from the experiment. Do the populations all contain the same number of different colored beads, or do they vary? Remember, these populations all came from the same exact parent population.
Form a conclusion: Most likely, the five resulting populations will differ quite dramatically. This is because natural disasters are not selective—they kill and spare individuals at random. Now think about how this might affect a real population. What happens when a hurricane hits the Mississippi Gulf Coast? How do the seabirds that live on the beach fare?
Gene Flow
Lecture Video: Gene flow
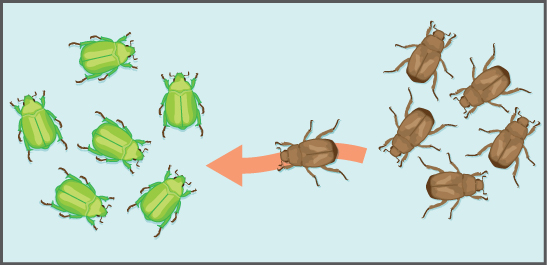
Another important evolutionary force is gene flow: the flow of alleles in and out of a population due to the migration of individuals or gametes (Figure 3). While some populations are fairly stable, others experience more flux. Many plants, for example, send their pollen far and wide, by wind or by bird, to pollinate other populations of the same species some distance away. Even a population that may initially appear to be stable, such as a pride of lions, can experience its fair share of immigration and emigration as developing males leave their mothers to seek out a new pride with genetically unrelated females. This variable flow of individuals in and out of the group not only changes the population’s gene structure, but it can also introduce new genetic variation to populations in different geological locations and habitats.
Mutation
Mutations are changes to an organism’s DNA and are an important driver of diversity in populations. Species evolve because of mutations accumulating over time. The appearance of new mutations is the most common way to introduce novel genotypic and phenotypic variance. Some mutations are unfavorable or harmful and are quickly eliminated from the population by natural selection. Others are beneficial and will spread through the population. Whether or not a mutation is beneficial or harmful is determined by whether it helps an organism survive to sexual maturity and reproduce. Some mutations do not do anything and can linger, unaffected by natural selection, in the genome. Some can have a dramatic effect on a gene and the resulting phenotype.
Lecture Video: Mutation and novel genetic variation.
Non-random Mating
If individuals non-randomly mate with their peers, the result can be a changing population. There are many reasons non-random mating occurs. One reason is simple mate choice. For example, female peahens may prefer peacocks with bigger, brighter tails. Natural selection picks traits that lead to more mating selections for an individual. One common form of mate choice, called assortative mating, is an individual’s preference to mate with partners who are phenotypically similar to themselves.
Another cause of nonrandom mating is physical location. This is especially true in large populations spread over vast geographic distances where not all individuals will have equal access to one another. Some might be miles apart through woods or over rough terrain, while others might live immediately nearby.
Inbreeding is another form of nonrandom mating that occurs when individuals mate with close relatives. Inbreeding is more common in small populations where potential mates are limited or in populations where individuals do not disperse widely from their birthplace. While mating with relatives can increase the likelihood of offspring inheriting advantageous traits shared within a family, it also raises the probability that harmful recessive alleles will be expressed. This can lead to inbreeding depression, a reduction in the population’s overall fitness due to an increase in genetic disorders and a decrease in genetic diversity. Inbreeding can make populations more susceptible to diseases and less adaptable to environmental changes. Understanding the impact of inbreeding is crucial, especially in conservation biology, where maintaining genetic diversity is essential for the survival of endangered species.
Lecture Video: Non-random Mating, inbreeding, and assortative mating.
Natural Selection
Natural selection is the process by which individuals with certain heritable traits tend to survive and reproduce more successfully than others, leading to a change in the frequency of those traits in the population.
Natural selection operates on phenotypes, the observable characteristics of organisms resulting from the interaction of their genotypes (genetic makeup) with the environment. While selection acts directly on phenotypes, it indirectly influences the underlying genotypes and allele frequencies within the population. The way alleles interact to produce a phenotype significantly affects how selection impacts genotype and allele frequencies.
Fitness in an evolutionary context refers to an individual’s genetic contribution to the next generation. It is measured by the number of viable offspring an individual produces compared to others in the population. Natural selection increases the average fitness of a population by favoring alleles that enhance survival and reproduction.
Although natural selection acts on phenotypes, the effects on genotype and allele frequencies depend on how alleles interact to produce those phenotypes. The relationship between alleles can be characterized by patterns of dominance, codominance, incomplete dominance, and overdominance, which influence how selection alters genetic variation.
In cases of complete dominance, one allele (the dominant allele) masks the effect of the other allele (the recessive allele) in heterozygous individuals. For instance:
- Dominant allele (A): Expressed in both AA and Aa genotypes.
- Recessive allele (a): Expressed only in the aa genotype.
When selection favors or disfavors a dominant or recessive phenotype, the impact on allele frequencies differs:
- Selection against a dominant allele: If the dominant phenotype is disadvantageous, both AA and Aa individuals are selected against because they express the dominant trait. This can lead to a rapid decrease in the frequency of the dominant allele since it’s exposed to selection in both homozygous and heterozygous forms.
- Selection against a recessive allele: If the recessive phenotype is disadvantageous, only aa individuals are selected against. The recessive allele remains hidden in heterozygous Aa individuals, allowing it to persist in the population. This makes it more challenging to eliminate the recessive allele entirely.
In cases of where the heterozygous genotype results in a different phenotypes than either homozygote, a pattern of overdominance or underdominance can occur:
- Overdominance (Heterozygote Advantage): The heterozygote genotype has a higher relative fitness than either homozygote. This can maintain genetic variation in the population because both alleles are preserved. The classic example is the sickle cell in humans, which is discussed further below.
- Underdominance: The heterozygote has lower fitness than both homozygotes. This can lead to disruptive selection, potentially resulting in one allele becoming fixed while the other is lost, depending on initial allele frequencies.
Population geneticists use mathematical models to predict how allele frequencies change under natural selection. For these models, population genetics often use a measure of relative fitness (w). Relative fitness refers to the reproductive success of a genotype or allele compared to others in the population. It quantifies how well individuals with a particular genotype contribute to the next generation relative to other genotypes. A genotype with higher relative fitness will increase in frequency over time, while those with lower fitness may decrease.
For example, if individuals with genotype AA produce more viable offspring than those with genotypes Aa or aa, then AA has a higher relative fitness. The differences in relative fitness among genotypes drive the changes in allele frequencies through natural selection.
Relative fitness (w) is typically scaled from 0 and 1 by dividing each genotypes’ fitness by the highest value.
| Genotype | Fitness | relative fitness (w) |
| AA | 100% survival rate (if AA is the most fit genotype) | wAA = 100/100 = 1.0 |
| Aa | 50% | wAa = 50/100 = 0.5 |
| aa | 20% | waa = 20/100 = 0.2 |
Selection coefficients (s) represent the reduction in fitness:
| Genotype | relative fitness (w) | Selection coefficients (s) |
| AA | 1.0 | s1 = 1-wAA = 0 (no selection) |
| Aa | 0.5 | s2 = 1-wAa = 0.5 |
| aa | 0.2 | s3 = 1-waa = 0.8 |
MATHMATICAL CONNECTION
See the following page for more on how evolutionary biologist calculate relative fitness and use it measure the strength of selection.
Examples of Selection and Allele Interactions
Sickle Cell Anemia (Overdominance)
One of the most well-known examples illustrating how allele interactions affect genotype frequencies under natural selection is the relationship between sickle cell anemia and malaria resistance in humans.
In regions where malaria is endemic, such as parts of sub-Saharan Africa, the Middle East, and South Asia, the presence of the sickle cell allele (HbS) influences survival rates. Malaria is caused by the Plasmodium parasite, which infects red blood cells. The genotypes and their associated phenotypes are as follows:
- HbA/HbS: Susceptible to malaria.
- HbA/HbS: Resistant to malaria and generally healthy.
- HbS/HbS: Suffer from sickle cell disease.
In regions where malaria is prevalent, individuals heterozygous for the sickle cell allele (HbA/HbS) have a survival advantage.
This scenario demonstrates heterozygote advantage (overdominance), where the heterozygous genotype (HbA/HbS) has a higher relative fitness than either homozygous genotype. In malaria-endemic regions, natural selection favors the heterozygotes because they have resistance to malaria without the severe disadvantages of sickle cell disease.
As a result, both the normal allele (HbA) and the sickle cell allele (HbS) are maintained in the population. The HbS allele persists at higher frequencies in these regions than would be expected if only considering its deleterious effects in homozygotes. This balance illustrates how allele interactions and environmental factors influence genotype frequencies under natural selection (Fig. 4).
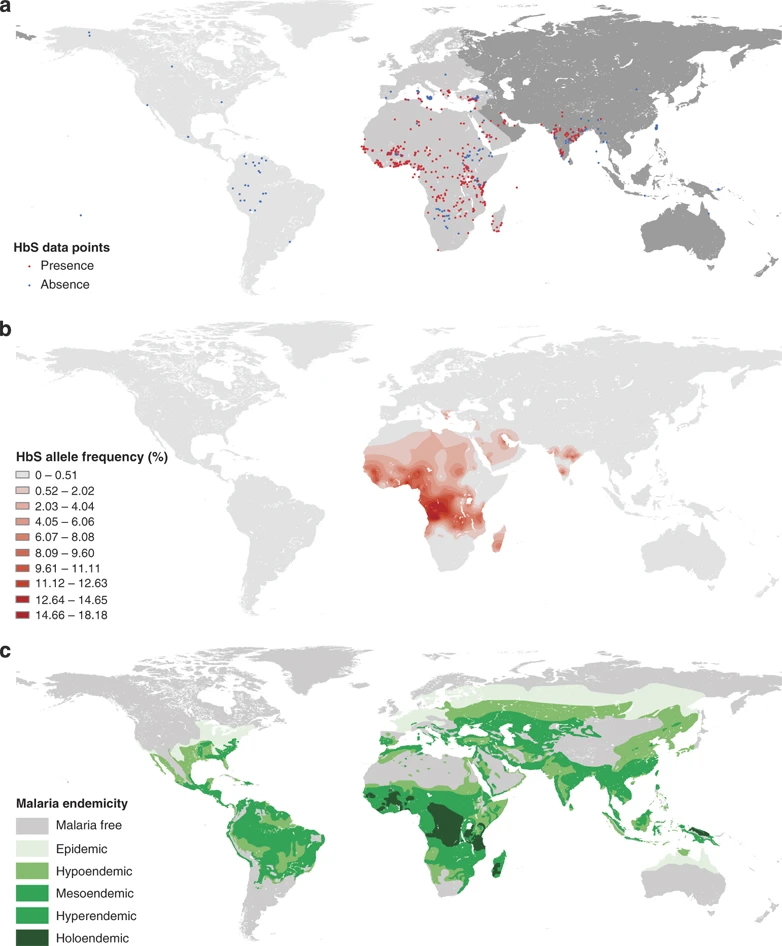
(b) A map showing the frequency of the HbS allele using a color gradient, with higher frequencies shown in darker shades.
(c) A historical map of malaria prevalence around the world. The map categorizes regions based on how common malaria is, ranging from malaria-free areas to those with high levels of the parasite in children. From: Piel et al. Nat Commun 1, 104 (2010). https://doi.org/10.1038/ncomms1104
Lethal Recessive Alleles
Alleles that are lethal when homozygous recessive (aa) can persist in a population because heterozygous individuals (Aa) are carriers without expressing the lethal phenotype. Selection reduces the frequency of the recessive allele slowly since it’s shielded from selection in heterozygotes.
Lecture video: Natural selection
Environmental Variance
Genes are not the only players involved in determining population variation. Other factors, such as the environment (Figure 5) also influence phenotypes. A beachgoer is likely to have darker skin than a city dweller, for example, due to regular exposure to the sun, an environmental factor. For some species, the environment determines some major characteristics, such as gender. For example, some turtles and other reptiles have temperature-dependent sex determination (TSD). TSD means that individuals develop into males if their eggs are incubated within a certain temperature range, or females at a different temperature range.

Geographic separation between populations can lead to differences in the phenotypic variation between those populations. We see such geographical variation between most populations and it can be significant. We can observe one type of geographic variation, a cline, as given species’ populations vary gradually across an ecological gradient. Species of warm-blooded animals, for example, tend to have larger bodies in the cooler climates closer to the earth’s poles, allowing them to better conserve heat. This is a latitudinal cline. Alternatively, flowering plants tend to bloom at different times depending on where they are along a mountain slope. This is an altitudinal cline.
If there is gene flow between the populations, the individuals will likely show gradual differences in phenotype along the cline. Restricted gene flow, alternatively can lead to abrupt differences, even speciation.
Footnotes
- 1 A. J. Tipping et al., “Molecular and Genealogical Evidence for a Founder Effect in Fanconi Anemia Families of the Afrikaner Population of South Africa,” PNAS 98, no. 10 (2001): 5734-5739, doi: 10.1073/pnas.091402398.
Glossary
- assortative mating
- when individuals tend to mate with those who are phenotypically similar to themselves
- bottleneck effect
- magnification of genetic drift as a result of natural events or catastrophes
- cline
- gradual geographic variation across an ecological gradient
- gene flow
- flow of alleles in and out of a population due to the individual or gamete migration
- genetic drift
- effect of chance on a population’s gene pool
- genetic variance
- diversity of alleles and genotypes in a population
- geographical variation
- differences in the phenotypic variation between populations that are separated geographically
- heritability
- fraction of population variation that can be attributed to its genetic variance
- inbreeding
- mating of closely related individuals
- inbreeding depression
- increase in abnormalities and disease in inbreeding populations
- mutation
- a change in the DNA sequence of an organism
- nonrandom mating
- changes in a population’s gene pool due to mate choice or other forces that cause individuals to mate with certain phenotypes more than others
- population variation
- distribution of phenotypes in a population
- selective pressure
- environmental factor that causes one phenotype to be better than another
Access for free at https://openstax.org/books/biology-2e/pages/1-introduction

{kind=link}