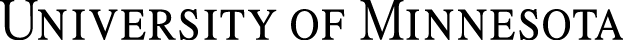
Appendix A: Supplemental Feline Notes
Esther Gallant and Christina Clarkson
FELINE ANATOMY NOTES
(for use as an appendix to Guide to the Dissection o f the Dog, 7th edition) Esther M. Gallant, PhD
Revised by Christina E. Clarkson, DVM, PhD
INTRODUCTION
This is not a comprehensive guide to cat anatomy, but rather a brief description of those aspects of feline anatomy which differ significantly from canine anatomy. Page and figure numbers for corresponding sections in Guide to the Dissection of the Dog, 7th Edition, (DG) are given in parentheses to facilitate comparisons between cat and dog morphology. Figures illustrating feline anatomy are not complete detailed representations but rather include only the structures of interest plus a few helpful landmarks for purposes of orientation. The asterisks (*) in the text indicate items of special clinical significance.
SUPERFICIAL STRUCTURES
Notice that cats have an average of 4 pair of mammary glands : 2 pair thoracic, 1 pair abdominal and 1 pair inguinal. (TVA). Some big cats have only the caudal pairs.
BONES OF THE FELINE THORACIC LIMB (DG. pp. 6-17)
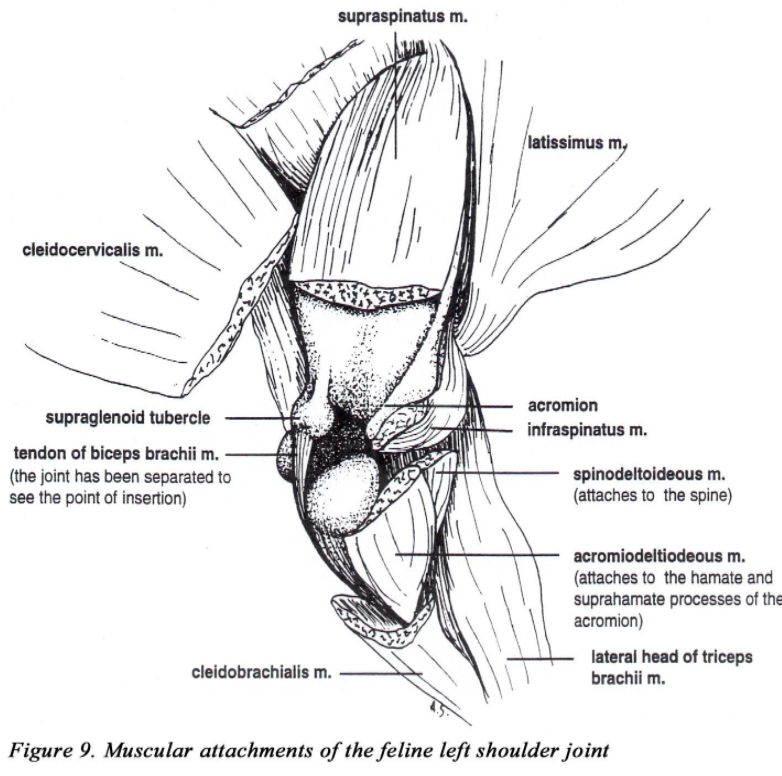
• The acromion of the scapula has two projections (hamate process and the suprahamate processes). The omotransversarius muscle (m.) takes origin from the acromion (suprahamate process) and inserts on the wing of the atlas (Figure 1A).
• A pronounced coracoid process projects medially from the supraglenoid tubercle. This is the origin of the coracobrachialis m. (Figure 1B).
• In the cat, a slender clavicle is present at the clavicular intersection of the brachiocephalicus m. (DG. p. 21). The ossified clavicle, which has no bony articulations, may be evident in radiographs (Figure 1C).
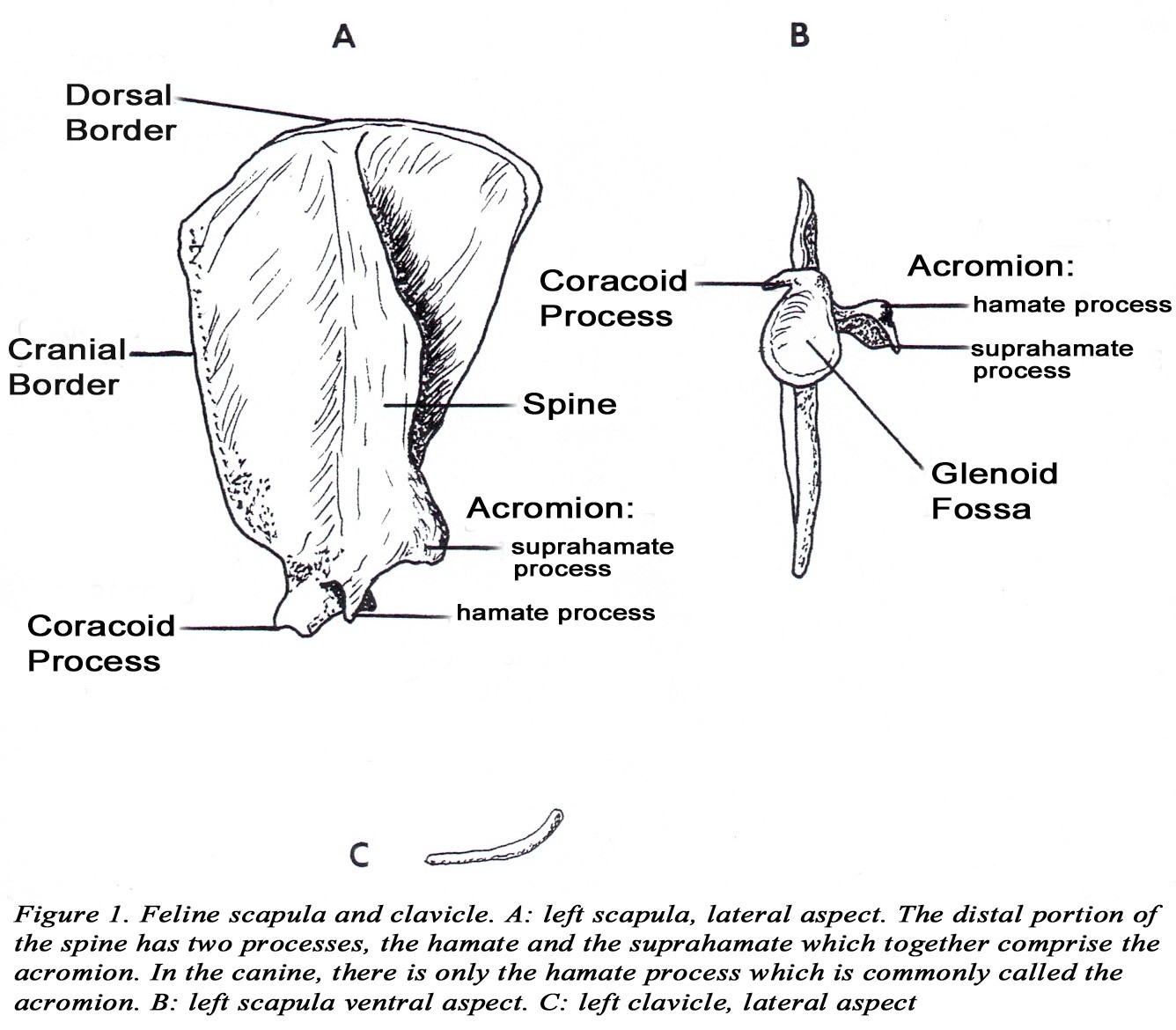
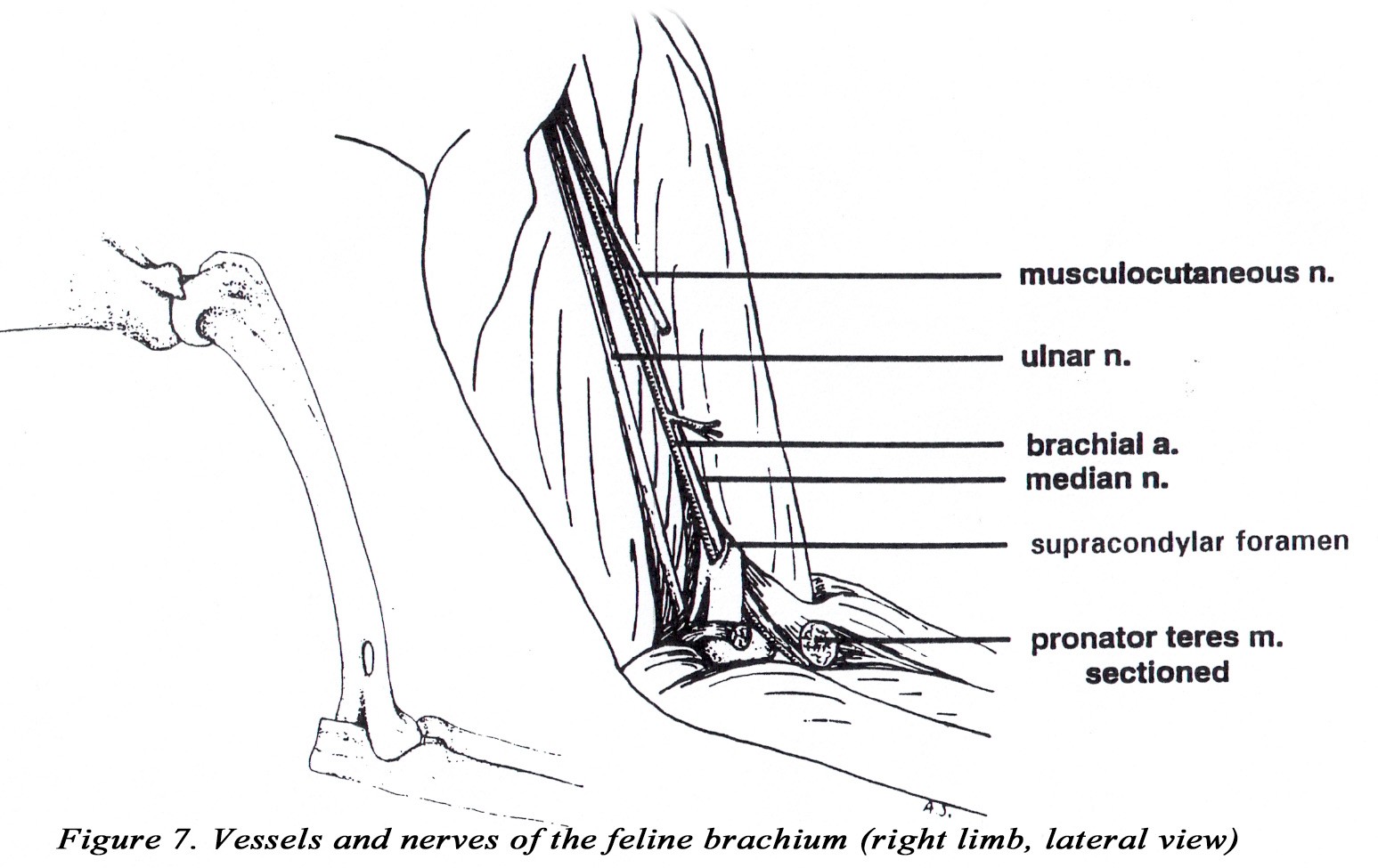
• A supracondylar foramen is present in the distal medial humerus. The brachial artery and median nerve pass through this foramen (Figure 2A). *This relationship can be important in the treatment of feline condyloid fractures of the humerus because of the possibility of damage to the artery and nerve where they pass through the foramen. Note: the dog has a supratrochlear foramen (nothing passes through it).
• Coronoid and radial fossae are present on the cranial side of the humoral condyle.
• The radial tuberosity projects caudally from the radius and is the site of insertion of the biceps brachii m. (Figure 2B).
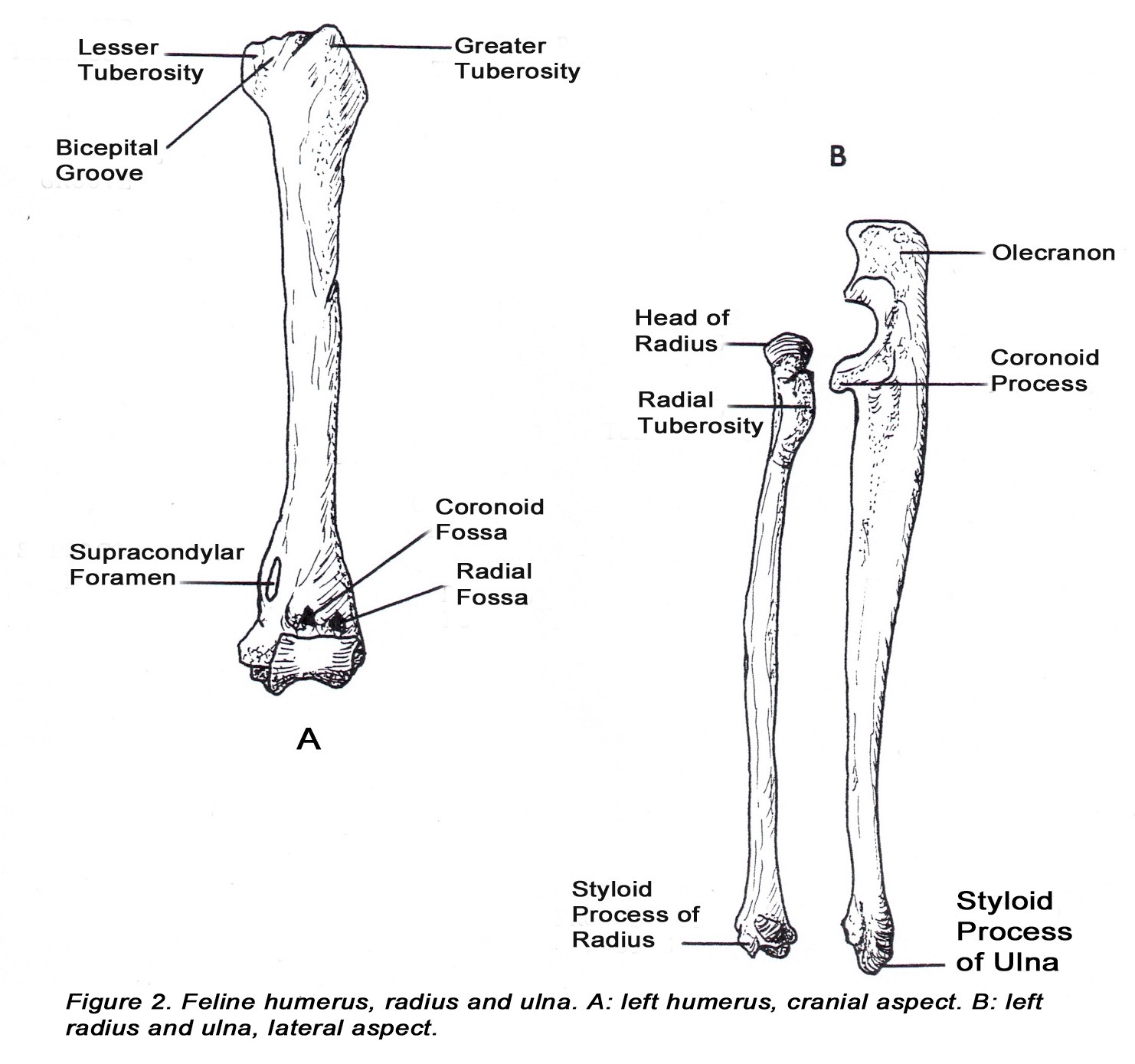
• The head of the middle phalanx is offset to the medial side of the distal phalanx of digits 2-5. This allows the distal phalanx to overlap the middle phalanx during the passive retraction of the distal phalanx.
• The distal phalanx has a deep concave caudal surface that articulates with the middle phalanx to form the distal interphalangeal joint (Figure 3C).
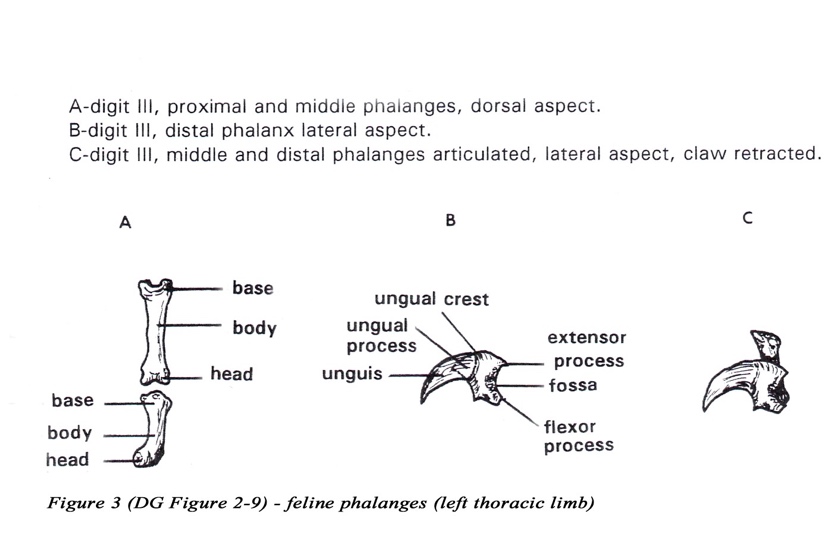
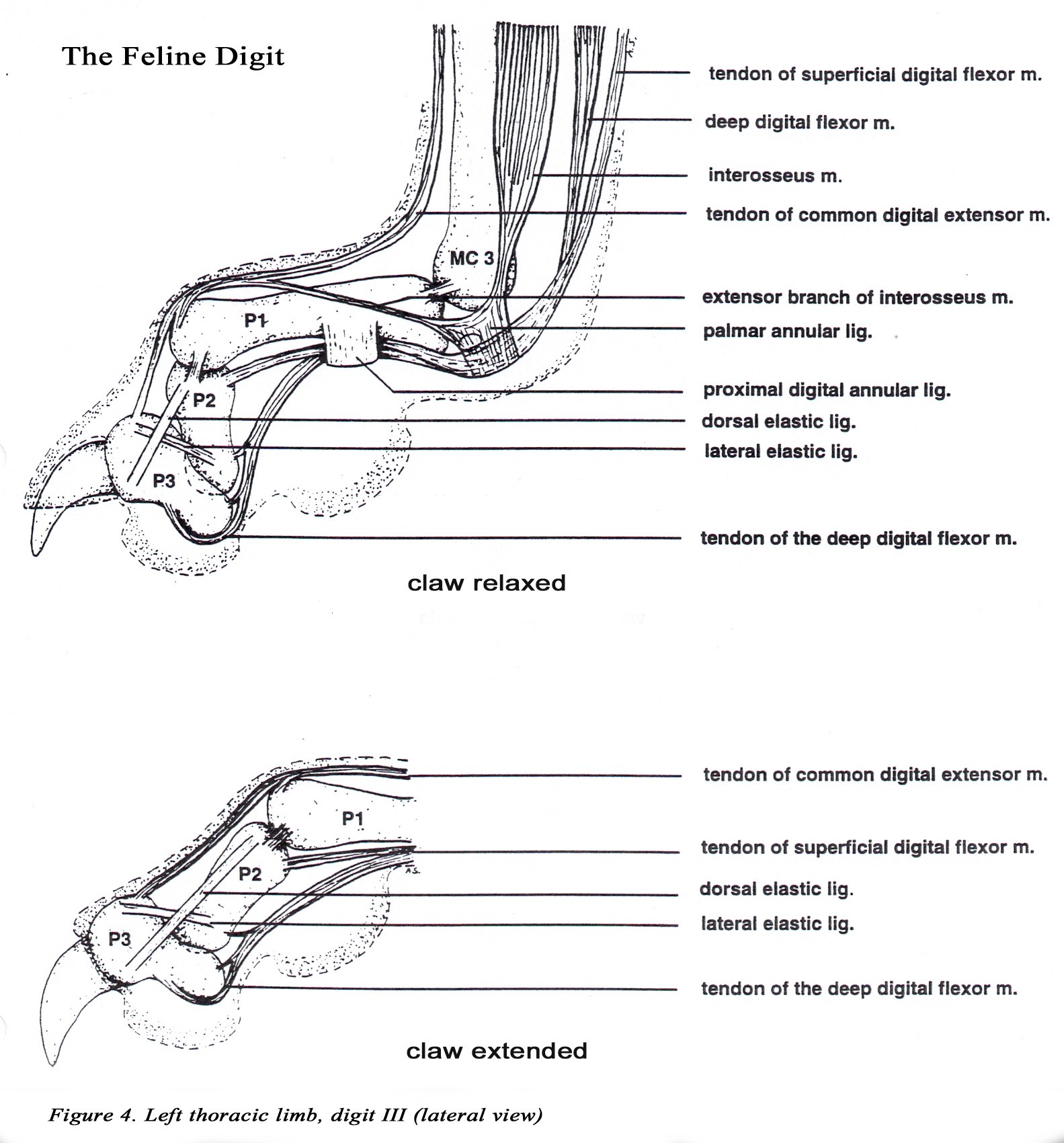
• Three elastic ligaments are responsible for passive retraction (dorsiflexion) of the claw. The most important elastic ligament, the lateral l igament , attaches to the extensor process of the distal phalanx and the lateral aspect of the head of the middle phalanx of digits 2-5. A pair of dorsal elastic ligaments extend from the sides of the extensor process to the base of the middle phalanx of digits 2- 5 (Figure 4).
• The dewclaw (digit 1, hallux) has proximal and distal phalanges only and cannot be retracted. The distal phalanx is similar to those of digits 2-5. The head of the proximal phalanx is not offset.
• The claw is protruded by contraction of the deep digital flexor m. which inserts on the flexor process of the distal phalanx. During claw protrusion, more proximal joints of the paw are fixed in position by action of the digital extensor mm. The ability to retract the claws keeps them sharp by preventing wear when not in use. All cats have retractable claws except the cheetah.
• *Declawing or onchyectomy may be performed through a partial amputation of the distal phalanx using a nail clipper, removing as much of the distal phalanx as possible. The deep digital flexor process should be all that remains in the joint. Another method involves disarticulation of the distal interphalangeal joint using a scalpel blade to dissect out the distal phalanx.
• Notice that the cat has an additional area of vibrissae (whiskers) in the caudal carpal area.
EXTRINSIC MUSCLES OF THE FELINE THORACIC LIMB (DG pp. 15-31)
• Note: cats do not have a nuchal ligament.
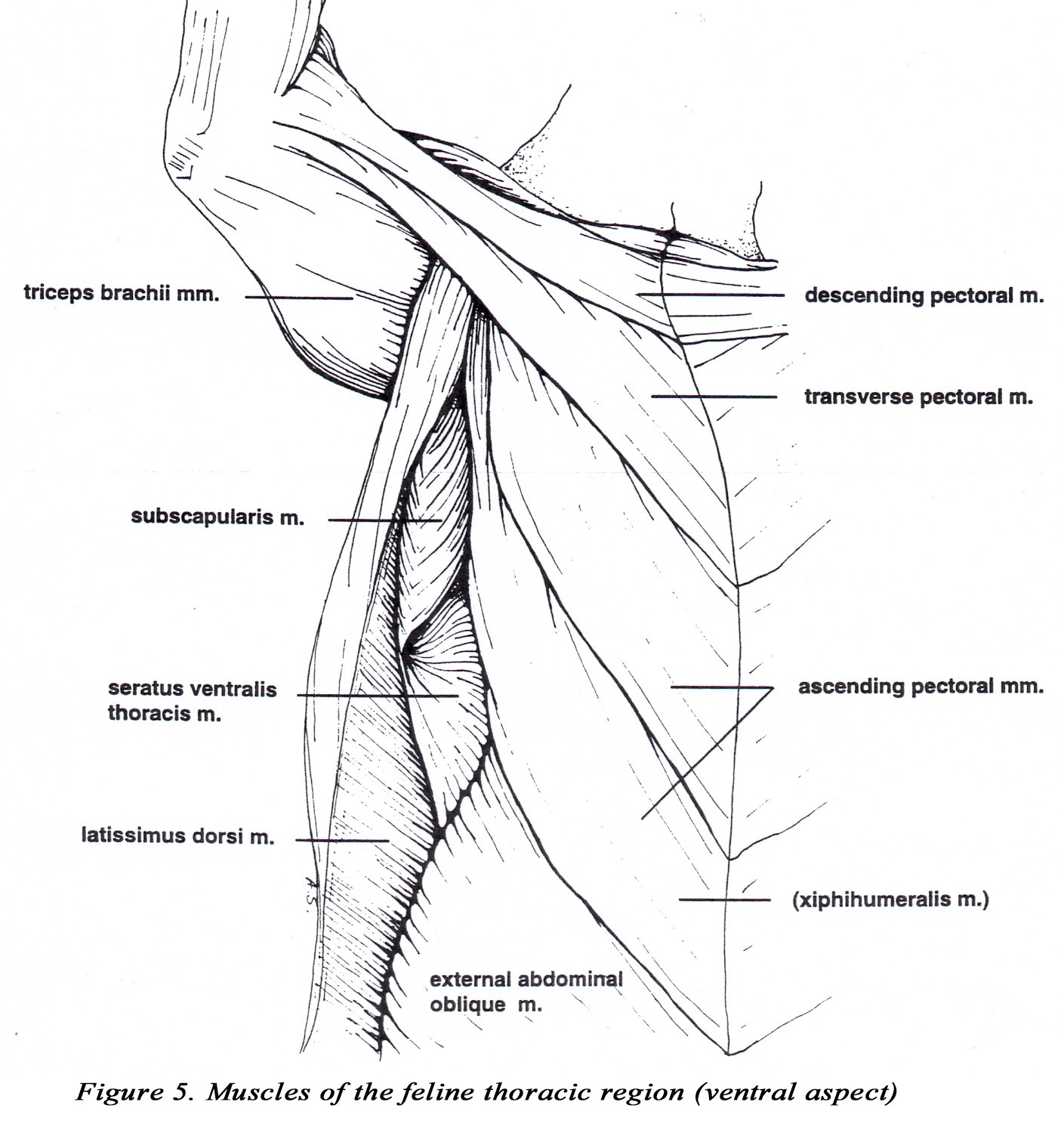
• The origin of the descending portion of the superficial pectoral m. is located more caudally than in the dog, and it extends to the medial fascia of the radius and ulna. A small tendinous portion attaches to the brachiocephalicus tendon. The transverse pectoral is relatively large and may appear fused to the deep pectoral m. A small fascicle from the deep pectoral m. also inserts on the medial fascia of the radius and ulna and may be difficult to separate from the superficial pectorals (Figure 5).
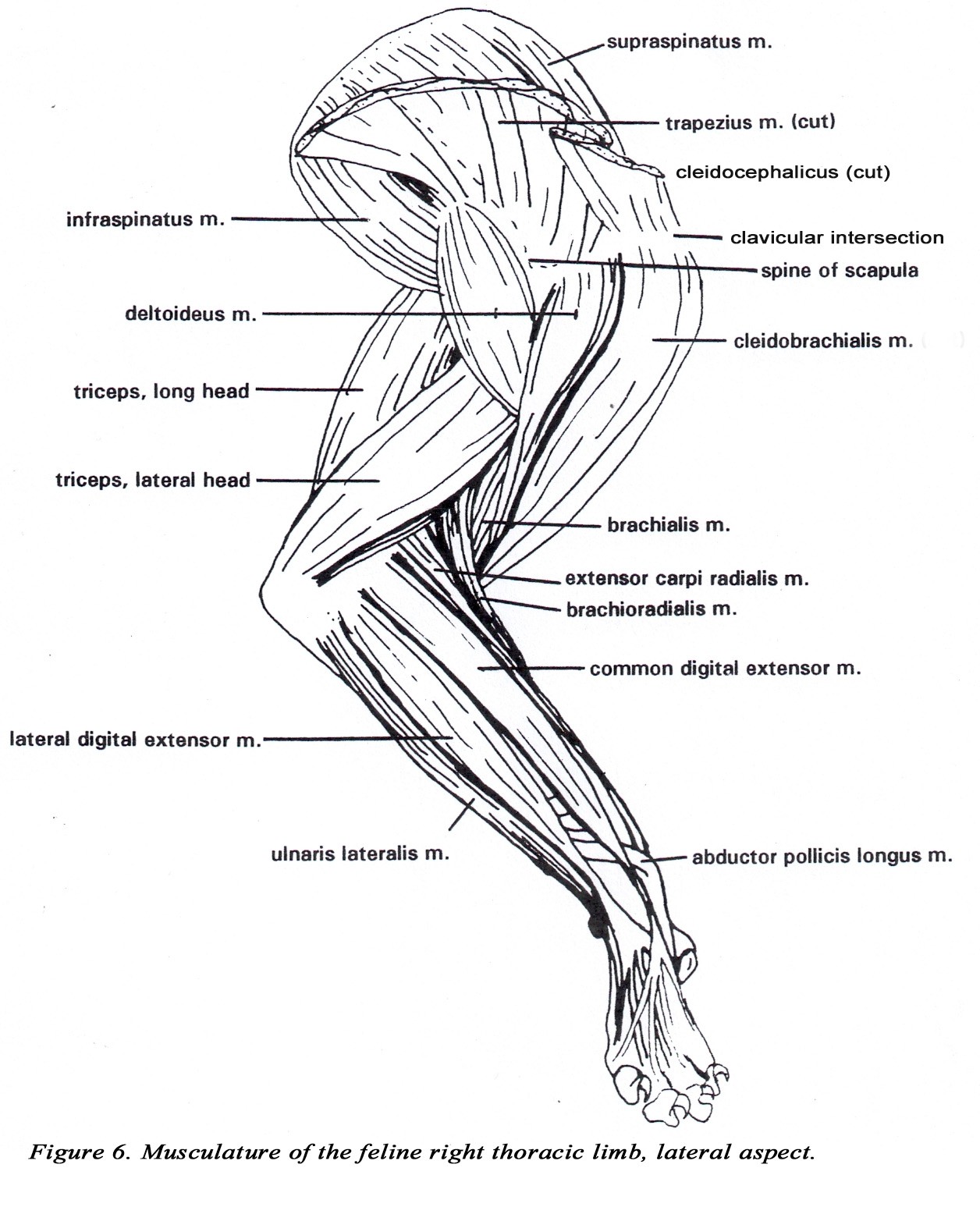
• A well defined osseous clavicle is present at the clavicular intersection of the brachiocephalicus m. , medial to the greater tubercle of the humerus. At the clavicular intersection, the muscle is much wider (relatively) than in the dog and may reach the ventral midline, overlapping and fusing with the superficial pectoral muscles. All three subdivisions of the brachiocephalicus m. are considered to arise from the clavicular intersection. The distal attachment of the cleidobrachialis m. is on the proximal medial ulna by a common tendon with the brachialis m. (Figure 6).
INTRINSIC MUSCLES OF THE FELINE THORACIC LIMB (DG p. 25-38)
• The two parts of the deltoideous m. (acromiodeltoideous and spinodeltoideous) are more separate and prominent in the cat.
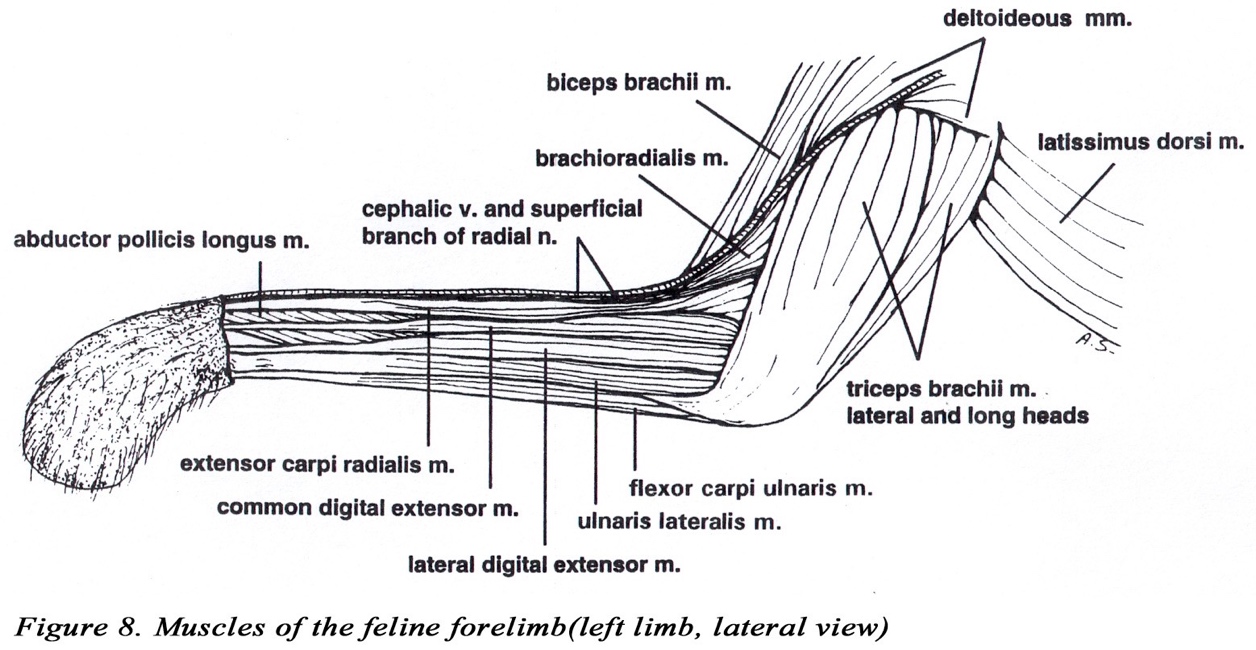
• The triceps brachii m. usually has four heads (long, lateral, medial, and accessory). The long head is usually not divided. The medial and accessory heads may appear fused, but the radial nerve can be observed passing between them. The medial head has an additional short part which arises from the distal medial side of the humerus, and inserts on the olecranon of the ulna. The median nerve and brachial artery pass through the supracondylar foramen which is covered by the short part of the medial head of the triceps brachii m. (Figures 2A & 7).
• The coracobrachialis tendon may be medial to the teres major m. and insert distal to it.
• The biceps brachii m. (as in the dog) has only one head which inserts by a single tendon on the radial tuberosity (Figures 2B & 8). The origin attaches to the supraglenoid tubercle of the scapula (Figure 9).
• The tendon of the brachialis m. along with the brachiocephalicus ( cleidobrachialis) tendon inserts by means of a common tendon on the proximal medial ulna.
• The brachioradialis m. is well developed in the cat. It originates from the proximal lateral humerus with the brachialis coursing distally with the superficial radial nerve and the cephalic vein to insert on the styloid process of the distal medial radius (Figures 6, 8,10).
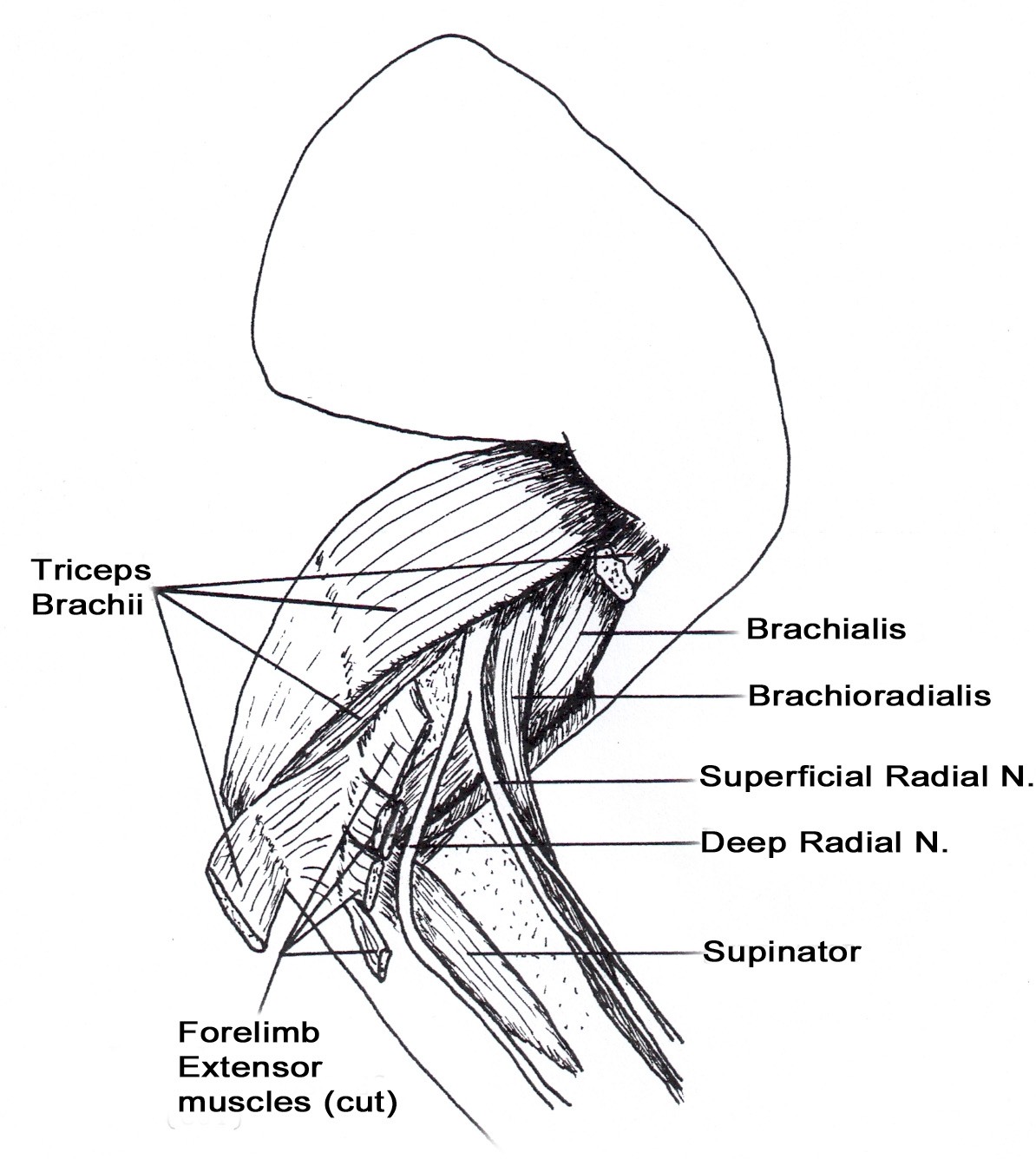
Figure 10. Proximal muscles of feline forelimb (right, lateral view)
• Abductor digiti I longus (a.k.a. abductor pollicis longus or extensor carpi obliquus)
• The extensor carpi radialis m. has a long and a short head. Both arise from the distal lateral humerus.
• The supinator m. lies deep to the deep radial nerve (Figure 10).
• The deep digital flexor m. has humoral, radial, and ulnar heads. The humeral head has three
parts. The ulnar and radial heads are much larger than in the dog.
• The pronator teres m. may appear fused to the radial head of the deep digital flexor.
• The ulnar nerve lies deep to the short part of the medial head of the triceps m. at the level of the olecranon (Figure 7).
• Review discussion of phalanges and retraction of claws on page 2-3.
BONES OF THE PELVIC LIMB (DG pp. 41-50)
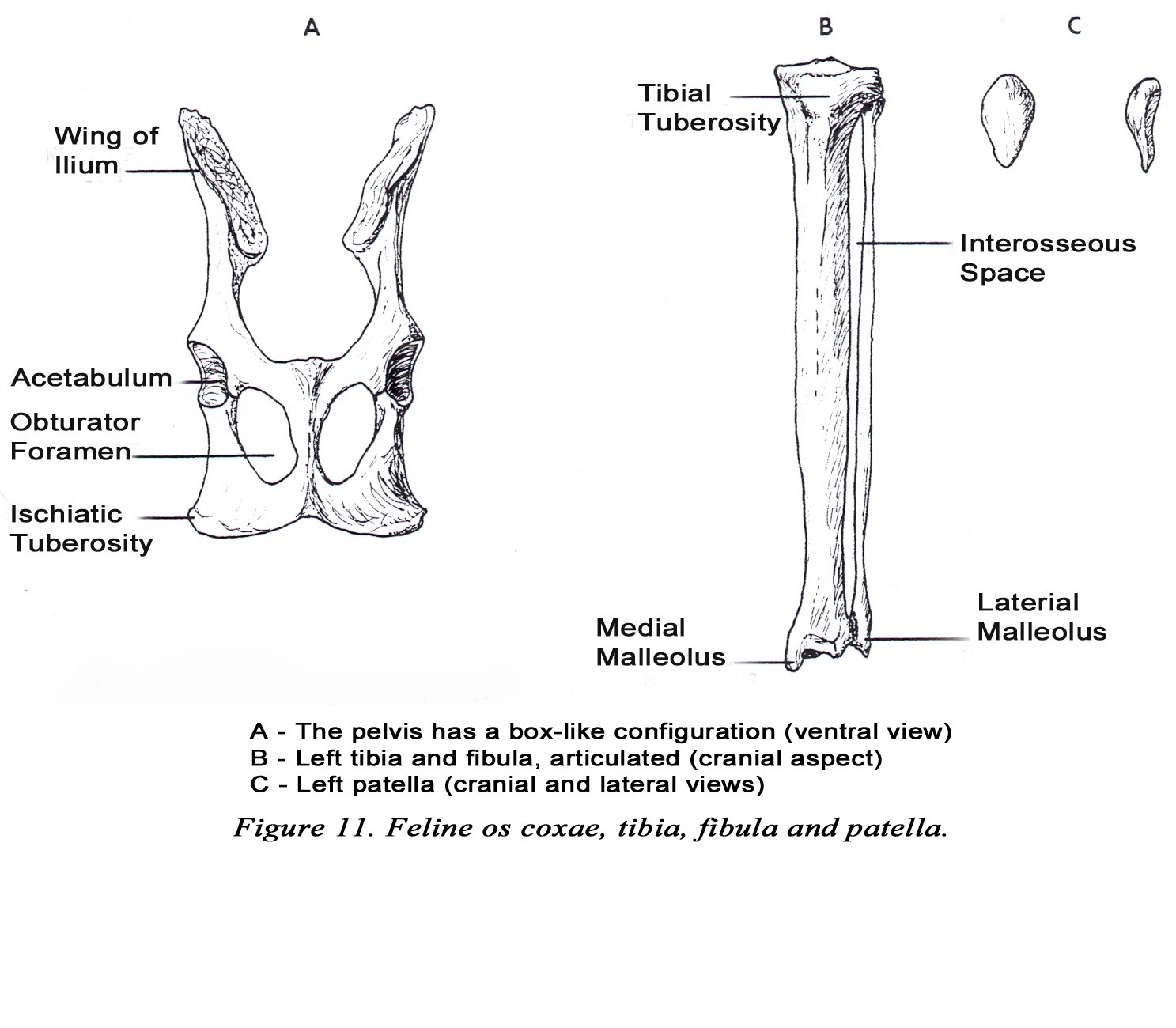
• In the ventral-dorsal view, the pelvis is nearly rectangular in shape (Figure 11A). The wing of the ilium and ischiatic tuberosity project laterally only slightly giving the pelvis a box-like appearance.
• The patella is pear-shaped in cats (Figure 11C). This is readily appreciated on radiographs.
• The interosseous space between the tibia and fibula is nearly the same width from the proximal to the distal articulation (Figure 11B).
MUSCLES OF THE FELINE PELVIC LIMB (DG pp. 50-69)
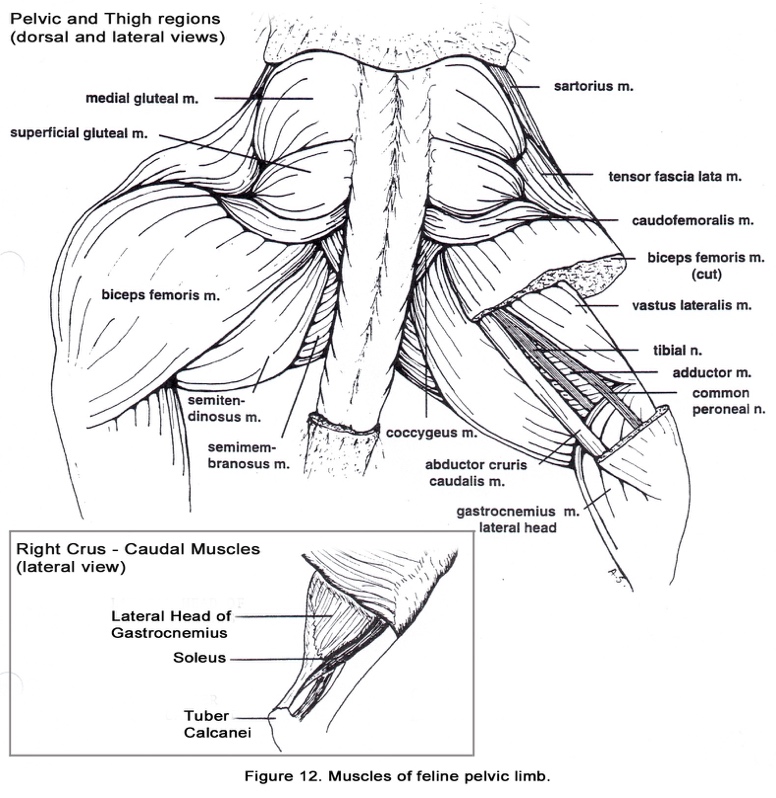
• The sacrotuberous ligament is absent in cats. Thus, the biceps femoris m. originates only from the ischiatic tuberosity (Figure 12).
• The gluteofemoralis (caudofemoralis m.) is found in cats adjacent to the biceps femoris m. (Figure 12). Its origin is on the transverse processes of the second and third caudal vertebrae. The muscle belly lies cranial to the biceps femoris m. and both muscles insert by means of the fascia lata on the patella, tibial tuberosity and cranial margin of the tibia.
• The sartorius m. usually appears to have only one belly which has the same insertion as the two bellies of the canine muscle. (Figures 12 & 13).
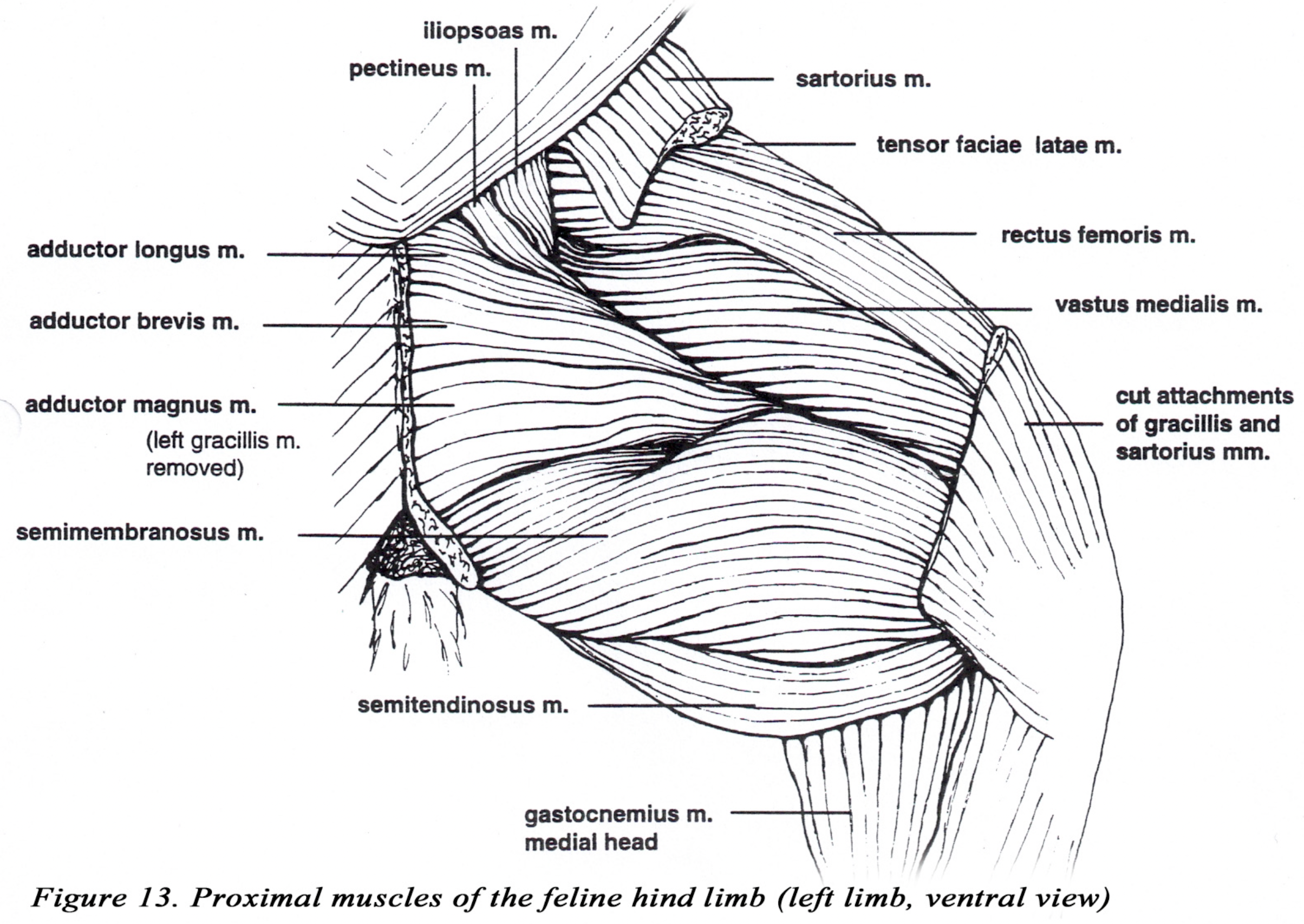
• The adductor longus m. is thin and adjacent to the pectineus m. which is also flattened in comparison to the canine muscle. (Figure 13).
• The vastus medialis and intermedius mm. are extensively fused and may also be partially fused to the vastus lateralis (Figures 12 & 13).
• The soleus m. is found in cats and many other mammals (Figure 12). It is deep to the lateral head of the gastrocnemius m.. The origin of the soleus is on the head of the fibula. The slender tendon of the soleus unites with the gastrocnemius tendon to form a major part of the common calcanean tendon. Notice the color of this muscle. An increase in mitochondria and myoglobin is responsible for the darker color of this fatigue-resistant muscle.
• The caudal tibial m. is very small in the dog (DG Fig. 2-55). In the cat, the caudal tibial m. is similar in size to the medial digital flexor and lies between it and the lateral digital flexor. As in the dog, the muscle arises from the head of the fibula and the caudal surface of the tibia. Its slender tendon parallels the medial digital flexor tendon in the distal crus and inserts on the medial plantar surface of the tarsus. It is an extensor of the hock.
MUSCLES OF THE TRUNK AND NECK (DG pp. 81-89)
• The serratus dorsalis m. (especially the pars caudalis) is more extensive in the cat (DG pp.82- 83).
• The biventer cervicis and the complexus mm. are generally difficult to separate. The longissimus capitus is relatively smaller than in the dog. The splenius and longissimus capitus mm. may be fused on their margins cranially. (DG p. 88)
• The nuchal ligament and supraspinous ligament are absent in the cat (DG p. 88). This allows greater flexibility of the vertebral column.
• The mandibular salivary gland is caudal to the maxillary vein and the mandibular lymph nodes are on either side of the linguofacial vein (DG p. 95).
• The omobrachial vein in the cat is usually absent. (DG pp. 96, 130)
THORAX
The thoracic duct enters the thorax caudally on the left lateral side of the aorta and continues cranially in this location. (DG pp. 105-106). The duct may be double for all or part of its length.
GASTROINTESTINAL
• The quadrate and right medial lobes of the l iver are fused. On the visceral surface, the fossa for the gall bladder marks the boundary between the quadrate lobe medially, and the right medial lobe laterally. (DG p. 149).
• The pancreatic duct to the major duodenal papilla is large. The accessory pancreatic duct is small and only present about 20% of the time. (DG p. 155)
• The cecum is comma shaped and lacks a cecocolic sphincter. (DG p. 155)
• The ascending colon is very short. (DG p. 155)
KIDNEY (DG pp. 155-157)
The feline kidney is true unipyramidal. Subcapsular stellate and interlobar veins are prominent on the surface of the kidney. The capsule may be quite tough.
FEMALE REPRODUCTIVE SYSTEM
• The round l igament of the uterus appears to originate from the mesometrium 1-2 cm. caudal to the ovary. (DG p. 158)
• The mesosalpinx envelopes approximately half of the ovary. (DG p. 158)
• Ovarian veins are not torturous.
ABDOMINAL ARTERIES (DG pp. 163-169)
The arterial supply to the abdomen is similar to the dog, with numerous variations. The arteries should be identified according to the region the vessel supplies and not by the branching pattern you observe.
PARANAL SINUSES ( ANAL SACS) (DG pp. 177-179)
The anal sacs of the cat have associated modified apocrine sweat glands and sebaceous glands. In the dog, sebaceous glands are not associated with the anal sacs.
MALE REPRODUCTIVE SYSTEM (DG pp. 180-183)
• The scrotum is immediately ventral to the anus and covered by hair.
• The cremaster m. typically absent or very small. An additional muscle, the levator scroti , originates from the external anal sphincter and inserts on the scrotal septum.
• A pair of bulbourethral glands are located caudal to the prostate. They are immediately proximal to the root of the penis and adjacent to the ischiocavernosus m. at the level of the ischial arch (Figure 14).
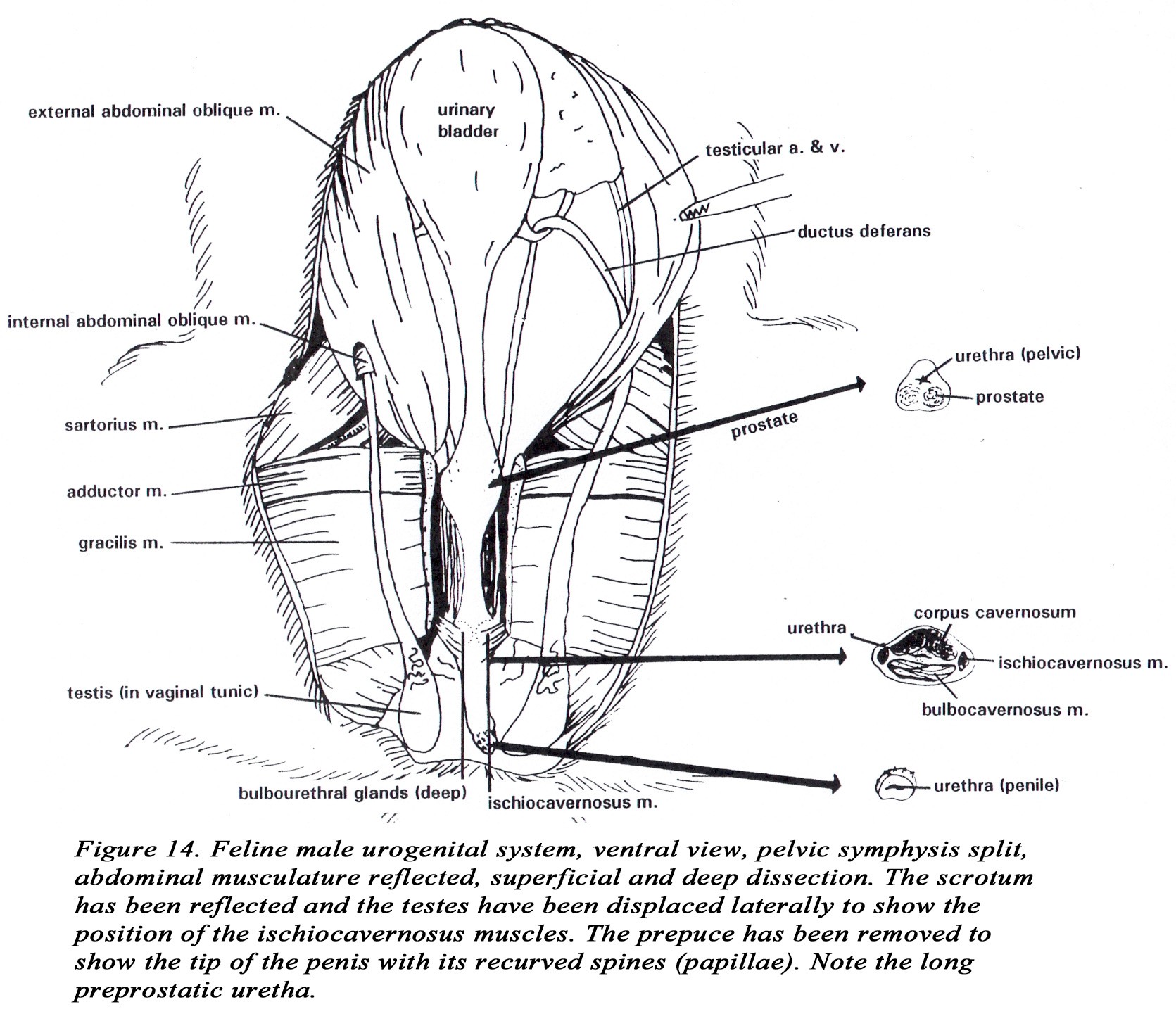
• The penis is short, directed caudally, and the glans is covered with curved papillae (spines). The glans penis is short and not differentiated into a pars bulbus and a pars longus. Otherwise, the structure of the penis is similar to the dog. Because of the caudal direction of the penis the dorsal artery and nerve are located ventrally.
• The os penis is small and may not be ossified.
• A long preprostatic urethra is present in the cat.
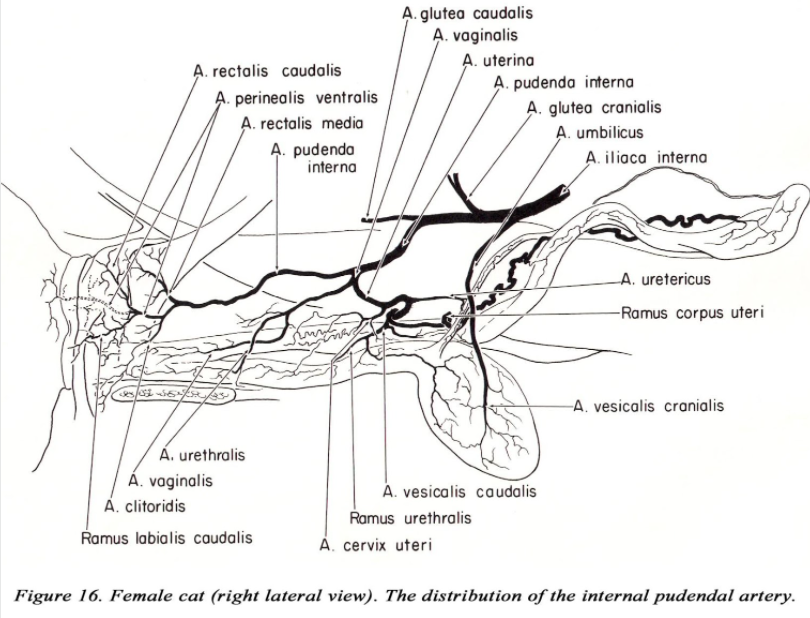
PELVIC ARTERIES (DG pp. 173-175)
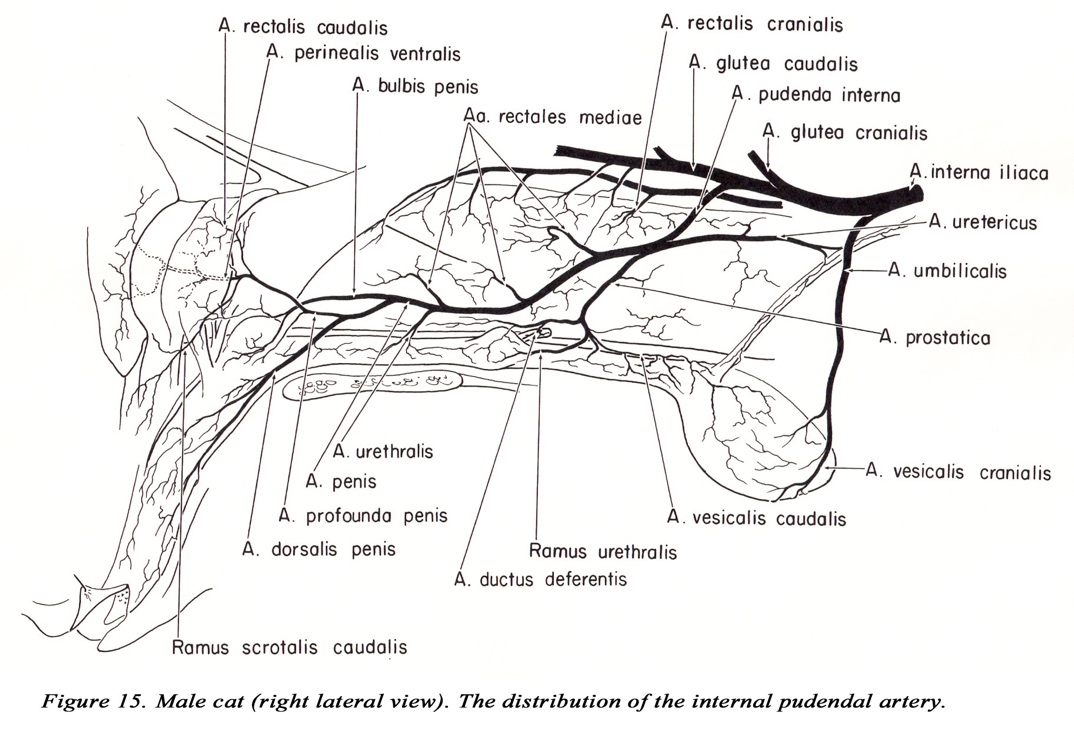
The branching of the internal i l iac artery varies considerably in the cat. As usual, identify the arteries by the area they supply. The first branch of the internal iliac is the umbilical a. The internal pudendal, caudal gluteal and iliolumbar aa. often arise at the same point, or the iliolumbar may arise near the umbilical a. The iliolumbar passes directly cranial to the shaft of the ilium. The caudal gluteal a. (after giving off the cranial gluteal a.) is small and may not extend to the caudal thigh musculature.
The pudendoepigastric trunk gives off an additional branch within the abdominal cavity, the middle vesical artery to the bladder. The internal pudendal artery in the cat remains within the pelvic canal throughout its course (in the dog the internal pudendal artery gives off visceral branches then passes obliquely across the greater ischiatic notch to become lateral to the coccygeus m. – DG p. 175).
HEAD
• The bony orbit is nearly complete. The frontal process of the zygomatic bone and the zygomatic process of the frontal bone are connected by a short orbital ligament to complete the orbit (DG p. 211).
• Cats lack an alar canal (DG p. 212). There is no trigeminal canal in the petrosal part of the temporal bone (DG p. 220-221).
• Dental formula for permanent teeth: (DG pp. 218-219)
| I | C | Pm | M | |||
| Upper | 3 | 1 | 3 | 1 | = 8 | |
| } x 2 = 30 total | ||||||
| Lower | 3 | 1 | 2 | 1 | = 7 |
• Filiform papillae of the cat’s tongue are large and directed caudally (DG p. 230). They serve as an aid in grooming.
• The buccal ( molar) gland is an additional salivary gland in cats located in the ventral buccal wall medial to the lower molar. Magnification is necessary to observe the orifice of the duct near the lower molar. The parotid duct enters near the upper molar (DG p. 233). The zygomatic salivary gland is located further ventral to the zygomatic arch than in the dog (DG p. 233)
• The laryngeal ventricles are absent from the larynx of the cat resulting in vocal ridges instead of vocal folds. The arytenoid cartilage lacks both corniculate and cuneiform processes and the vestibular fold is absent (DG pp. 235-236).
*Purring is under voluntary control by a rhythmic contraction of laryngeal muscles. The recurrent laryngeal nerve innervates these muscles. Inspiration and expiration phases are nearly equal while purring. The laryngeal muscles (cricoarytenoideus lateralis and thyroarytenoideus) contract producing stenosis at the glottis and causing a change in trans-glottal pressure. When the glottis begins to open (cricoarytenoides dorsalis) the purring sound occurs. This cycle repeats about 25 times/minute resulting in the palpable and audible sound of purring. To auscultate (hear) the heart and lungs, briefly occlude the external nares.
• The free margins of the feline tracheal cartilages overlap dorsally. The trachealis m. is dorsal to these rings.
• When the feline iris contracts (miosis) the pupil appears as a line. This is due to the direction of orientation of the muscle.
• Cats do not have cilia (eyelashes) on either eyelid.
• The esophagus in the cat has striated (voluntary) muscle along only 2/3 of its length. Vomition in the cat is more difficult than in the dog.
• The internal carotid artery is atrophied in the adult cat (DG p. 249). Instead, the ascending pharyngeal artery (DG Figure 5-43 & 5-45) supplies the arterial circle of the brain via the distal remnant of the internal carotid artery. The carotid sinus is located at the base of the occipital and ascending pharyngeal arteries (DG p. 249).
ACKNOWLEDGEMENTS:
Figures 6, 10, 12b, 14 by Daina Rosen, ’87.
Figures 4, 5, 7, 8, 9, 12a 13, by Ann Silverness, ’97.
Figures 15 and 16 from William Martin (Thesis 1973)
SUPPLEMENTAL INFORMATION
The Cat Family
Order – Carnivora
Family – Felidae
Genus and species (37 species) Felis catus – domestic cat
Felis concolor – puma (a.k.a. cougar or mountain lion) Felis pardalis – ocelot
Lynx rufus – bobcat
Lynx canadensis – Canada lynx
Panthera leo – lion Panthera onca – jaguar Panthera pardus – leopard Panthera tigris – tiger
Acinonyx jubatus – cheetah
