Speciation
Speciation
What are species anyway, and how do new ones evolve?
Here, you can explore different ways to define a species and learn about the various processes through which speciation can occur. This section also addresses the topics of cospeciation — when two lineages split in concert with one another — and modes of speciation that are specific to plants.
Let’s start by defining a species.

Defining a species
A species is often defined as a group of individuals that actually or potentially interbreed in nature. In this sense, a species is the biggest gene pool possible under natural conditions.
For example, these happy face spiders look different, but since they can interbreed, they are considered the same species: Theridion grallator.

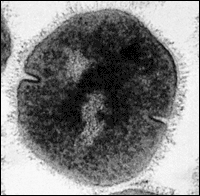 That definition of a species might seem cut and dried, but it is not — in nature, there are lots of places where it is difficult to apply this definition. For example, many bacteria reproduce mainly asexually. The bacterium shown at right is reproducing asexually, by binary fission. The definition of a species as a group of interbreeding individuals cannot be easily applied to organisms that reproduce only or mainly asexually.
That definition of a species might seem cut and dried, but it is not — in nature, there are lots of places where it is difficult to apply this definition. For example, many bacteria reproduce mainly asexually. The bacterium shown at right is reproducing asexually, by binary fission. The definition of a species as a group of interbreeding individuals cannot be easily applied to organisms that reproduce only or mainly asexually.
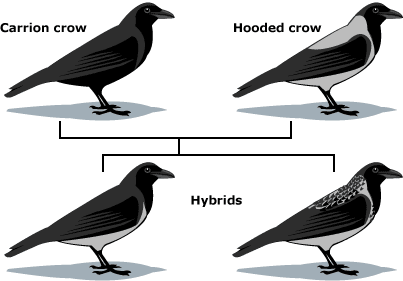
Also, many plants, and some animals, form hybrids in nature. Hooded crows and carrion crows look different, and largely mate within their own groups — but in some areas, they hybridize. Should they be considered the same species or separate species?
If two lineages of oak look quite different, but occasionally form hybrids with each other, should we count them as different species? There are lots of other places where the boundary of a species is blurred. It’s not so surprising that these blurry places exist — after all, the idea of a species is something that we humans invented for our own convenience!
Defining speciation
Speciation is a lineage-splitting event that produces two or more separate species. Imagine that you are looking at a tip of the tree of life that constitutes a species of fruit fly. Move down the phylogeny to where your fruit fly twig is connected to the rest of the tree. That branching point, and every other branching point on the tree, is a speciation event. At that point genetic changes resulted in two separate fruit fly lineages, where previously there had just been one lineage. But why and how did it happen?
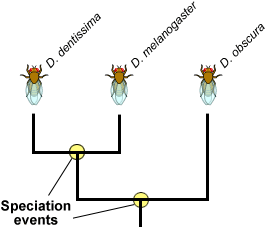
The branching points on this partial Drosophila phylogeny represent long past speciation events. Here is one scenario that exemplifies how speciation can happen:
- The scene: a population of wild fruit flies minding its own business on several bunches of rotting bananas, cheerfully laying their eggs in the mushy fruit…

- Disaster strikes: A hurricane washes the bananas and the immature fruit flies they contain out to sea. The banana bunch eventually washes up on an island off the coast of the mainland. The fruit flies mature and emerge from their slimy nursery onto the lonely island. The two portions of the population, mainland and island, are now too far apart for gene flow to unite them. At this point, speciation has not occurred — any fruit flies that got back to the mainland could mate and produce healthy offspring with the mainland flies.

- The populations diverge: Ecological conditions are slightly different on the island, and the island population evolves under different selective pressures and experiences different random events than the mainland population does. Morphology, food preferences, and courtship displays change over the course of many generations of natural selection.

- So we meet again: When another storm reintroduces the island flies to the mainland, they will not readily mate with the mainland flies since they’ve evolved different courtship behaviors. The few that do mate with the mainland flies, produce inviable eggs because of other genetic differences between the two populations. The lineage has split now that genes cannot flow between the populations.

This is a simplified model of speciation by geographic isolation, but it gives an idea of some of the processes that might be at work in speciation. In most real-life cases, we can only put together part of the story from the available evidence. However, the evidence that this sort of process does happen is strong.
Causes of speciation
Geographic isolation
In the fruit fly example, some fruit fly larvae were washed up on an island, and speciation started because populations were prevented from interbreeding by geographic isolation. Scientists think that geographic isolation is a common way for the process of speciation to begin: rivers change course, mountains rise, continents drift, organisms migrate, and what was once a continuous population is divided into two or more smaller populations.
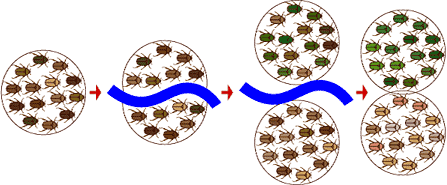
It doesn’t even need to be a physical barrier like a river that separates two or more groups of organisms — it might just be unfavorable habitat between the two populations that keeps them from mating with one another.
Reduction of gene flow
However, speciation might also happen in a population with no specific extrinsic barrier to gene flow. Imagine a situation in which a population extends over a broad geographic range, and mating throughout the population is not random. Individuals in the far west would have zero chance of mating with individuals in the far eastern end of the range. So we have reduced gene flow, but not total isolation. This may or may not be sufficient to cause speciation. Speciation would probably also require different selective pressures at opposite ends of the range, which would alter gene frequencies in groups at different ends of the range so much that they would not be able to mate if they were reunited.
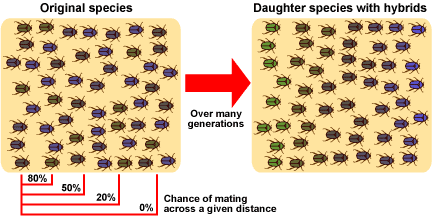
Even in the absence of a geographic barrier, reduced gene flow across a species’ range can encourage speciation.
Reproductive isolation
The environment may impose an external barrier to reproduction, such as a river or mountain range, between two incipient species but that external barrier alone will not make them separate, full-fledged species. Allopatry may start the process off, but the evolution of internal (i.e., genetically-based) barriers to gene flow is necessary for speciation to be complete. If internal barriers to gene flow do not evolve, individuals from the two parts of the population will freely interbreed if they come back into contact. Whatever genetic differences may have evolved will disappear as their genes mix back together. Speciation requires that the two incipient species be unable to produce viable offspring together or that they avoid mating with members of the other group.
Here are some of the barriers to gene flow that may contribute to speciation. They result from natural selection, sexual selection, or even genetic drift:
- The evolution of different mating location, mating time, or mating rituals:
Genetically-based changes to these aspects of mating could complete the process of reproductive isolation and speciation. For example, bowerbirds (shown below) construct elaborate bowers and decorate them with different colors in order to woo females. If two incipient species evolved differences in this mating ritual, it might permanently isolate them and complete the process of speciation.

Different species of bowerbird construct elaborate bowers and decorate them with different colors in order to woo females. The Satin bowerbird (left) builds a channel between upright sticks, and decorates with bright blue objects, while the MacGregor’s Bowerbird (right) builds a tall tower of sticks and decorates with bits of charcoal. Evolutionary changes in mating rituals, such as bower construction, can contribute to speciation. - Lack of “fit” between sexual organs:
Hard to imagine for us, but a big issue for insects with variably-shaped genitalia!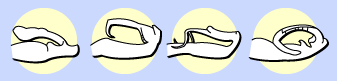
These damselfly penises illustrate just how complex insect genitalia may be. - Offspring inviability or sterility:
All that courting and mating is wasted if the offspring of matings between the two groups do not survive or cannot reproduce.
In our fruit-flies-in-rotten-bananas-in-a-hurricane example, allopatry kicked off the speciation process, but different selection pressures on the island caused the island population to diverge genetically from the mainland population.
What might have caused that to happen? Perhaps, different fruits were abundant on the island. The island population was selected to specialize on a particular type of fruit and evolved a different food preference from the mainland flies.
Could this small difference be a barrier to gene flow with the mainland flies? Yes, if the flies find mates by hanging out on preferred foods, then if they return to the mainland, they will not end up mating with mainland flies because of this different food preference. Gene flow would be greatly reduced; and once gene flow between the two species is stopped or reduced, larger genetic differences between the species can accumulate.

Evidence for speciation
Speciation in action?
In the summer of 1995, at least 15 iguanas survived Hurricane Marilyn on a raft of uprooted trees. They rode the high seas for a month before colonizing the Caribbean island, Anguilla. These few individuals were perhaps the first of their species, Iguana iguana, to reach the island. If there were other intrepid Iguana iguana colonizers of Anguilla, they died out before humans could record their presence.
 Evolutionary biologists would love to know what happens next: will the colonizing iguanas die out, will they survive and change only slightly, or will they become reproductively isolated from other Iguana iguana and become a new species? We could be watching the first steps of an allopatric speciation event, but in such a short time we can’t be sure.
Evolutionary biologists would love to know what happens next: will the colonizing iguanas die out, will they survive and change only slightly, or will they become reproductively isolated from other Iguana iguana and become a new species? We could be watching the first steps of an allopatric speciation event, but in such a short time we can’t be sure.
A plausible model
We have several plausible models of how speciation occurs — but of course, it’s hard for us to get an eye-witness account of a natural speciation event since most of these events happened in the distant past. We can figure out that speciation events happened and often when they happened, but it’s more difficult to figure out how they happened. However, we can use our models of speciation to make predictions and then check these predictions against our observations of the natural world and the outcomes of experiments. As an example, we’ll examine some evidence relevant to the allopatric speciation model.
Scientists have found a lot of evidence that is consistent with allopatric speciation being a common way that new species form:
- Geographic patterns: If allopatric speciation happens, we’d predict that populations of the same species in different geographic locations would be genetically different. There are abundant observations suggesting that this is often true. For example, many species exhibit regional “varieties” that are slightly different genetically and in appearance, as in the case of the Northern Spotted Owl and the Mexican Spotted Owl. Also, ring species are convincing examples of how genetic differences may arise through reduced gene flow and geographic distance.
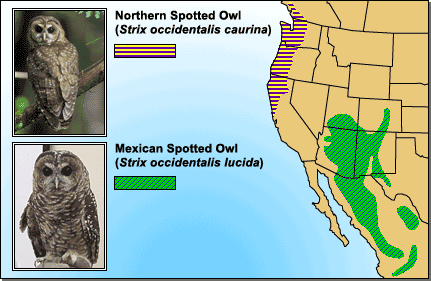
Spotted owl subspecies living in different geographic locations show some genetic and morphological differences. This observation is consistent with the idea that new species form through geographic isolation. - Experimental results: The first steps of speciation have been produced in several laboratory experiments involving “geographic” isolation. For example, Diane Dodd examined the effects of geographic isolation and selection on fruit flies. She took fruit flies from a single population and divided them into separate populations living in different cages to simulate geographic isolation. Half of the populations lived on maltose-based food, and the other populations lived on starch-based foods. After many generations, the flies were tested to see which flies they preferred to mate with. Dodd found that some reproductive isolation had occurred as a result of the geographic isolation and selection for different food sources in the two environments: “maltose flies” preferred other “maltose flies,” and “starch flies” preferred other “starch flies.” Although, we can’t be sure, these preference differences probably existed because selection for using different food sources also affected certain genes involved in reproductive behavior. This is the sort of result we’d expect, if allopatric speciation were a typical mode of speciation.
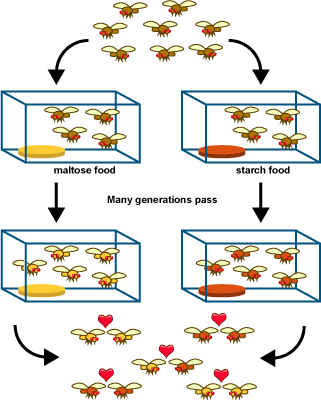
Diane Dodd’s fruit fly experiment suggests that isolating populations in different environments (e.g., with different food sources) can lead to the beginning of reproductive isolation. These results are consistent with the idea that geographic isolation is an important step of some speciation events.
Cospeciation
If the association between two species is very close, they may speciate in parallel. This is called cospeciation. It is especially likely to happen between parasites and their hosts.
To see how it works, imagine a species of louse living on a species of gopher. When the gophers get together to mate, the lice get an opportunity to switch gophers and perhaps mate with lice on another gopher. Gopher-switching allows genes to flow through the louse species.

Consider what happens to the lice if the gopher lineage splits into lineages A and B:
- Lice have few opportunities for gopher-switching, and lice on gopher lineage A don’t mate with lice living on gopher lineage B.
- This “geographic” isolation of the louse lineages may cause them to become reproductively isolated as well, and hence, separate species.
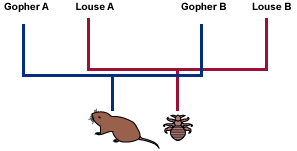
Evolutionary biologists can often tell when lineages have cospeciated because the parasite phylogeny will “mirror” the host phylogeny.
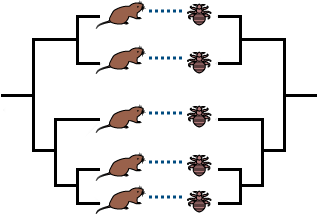
This example is somewhat idealized — rarely do scientists find hosts and parasites with exactly matching phylogenies. However, sometimes the phylogenies indicate that cospeciation did happen along with some host-switching.
Attribution
“Speciation.” Understanding Evolution. University of California Museum of Paleontology. 8 June 2020 <https://evolution.berkeley.edu/evolibrary/article/evo_40>.
“Defining a species.” Understanding Evolution. University of California Museum of Paleontology. 8 June 2020 <https://evolution.berkeley.edu/evolibrary/article/evo_41>.
“Defining speciation.” Understanding Evolution. University of California Museum of Paleontology. 8 June 2020 <https://evolution.berkeley.edu/evolibrary/article/evo_42>.
“Causes of speciation.” Understanding Evolution. University of California Museum of Paleontology. 8 June 2020 <https://evolution.berkeley.edu/evolibrary/article/evo_43>.
“Reproductive isolation.” Understanding Evolution. University of California Museum of Paleontology. 8 June 2020 <https://evolution.berkeley.edu/evolibrary/article/evo_44>.
“Evidence for speciation.” Understanding Evolution. University of California Museum of Paleontology. 8 June 2020 <https://evolution.berkeley.edu/evolibrary/article/evo_45>.
“Cospeciation.” Understanding Evolution. University of California Museum of Paleontology. 8 June 2020 <https://evolution.berkeley.edu/evolibrary/article/evo_46>.
All of the genes in a population. Any genes that could wind up in the same individual through sexual reproduction are in the same gene pool.
The movement of genes between populations. This may happen through the migration of organisms or the movement of gametes (such as pollen blown to a new location).
A group of organisms that is about to become a separate species from other, related individuals.
Speciation that depends on an external barrier to gene flow (such as geographic isolation) to begin or complete the process of speciation.
Speciation that depends on an external barrier to gene flow (such as geographic isolation) to begin or complete the process of speciation.
