98 Vertebrates
By the end of this section, you will be able to:
- Describe the difference between jawless and jawed fishes
- Explain the main characteristics of amphibians, reptiles, and birds
- Describe the derived characteristics in birds that facilitate flight
- Name and describe the distinguishing features of the three main groups of mammals
- Describe the derived features that distinguish primates from other animals
Vertebrates are among the most recognizable organisms of the animal kingdom (Figure 1). More than 62,000 vertebrate species have been identified. The vertebrate species now living represent only a small portion of the vertebrates that have existed. The best-known extinct vertebrates are the dinosaurs, a unique group of reptiles, reaching sizes not seen before or since in terrestrial animals. They were the dominant terrestrial animals for 150 million years, until they died out near the end of the Cretaceous period in a mass extinction. A great deal is known about the anatomy of the dinosaurs, given the preservation of their skeletal elements in the fossil record.

Fishes
Modern fishes include an estimated 31,000 species. Fishes were the earliest vertebrates, and jawless fishes were the earliest of these. Jawless fishes—the present day hagfishes and lampreys—have a distinct cranium and complex sense organs including eyes, distinguishing them from the invertebrate chordates. The jawed fishes evolved later and are extraordinarily diverse today. Fishes are active feeders, rather than sessile, suspension feeders.
Jawless Fishes
Jawless fishes are craniates (which includes all the chordate groups except the tunicates and lancelets) that represent an ancient vertebrate lineage that arose over one half-billion years ago. Some of the earliest jawless fishes were the ostracoderms (which translates as “shell-skin”). Ostracoderms, now extinct, were vertebrate fishes encased in bony armor, unlike present-day jawless fishes, which lack bone in their scales.
The clade Myxini includes 67 species of hagfishes. Hagfishes are eel-like scavengers that live on the ocean floor and feed on dead invertebrates, other fishes, and marine mammals (Figure 2a). Hagfishes are entirely marine and are found in oceans around the world except for the polar regions. A unique feature of these animals is the slime glands beneath the skin that are able to release an extraordinary amount of mucus through surface pores. This mucus may allow the hagfish to escape from the grip of predators. Hagfish are known to enter the bodies of dead or dying organisms to devour them from the inside.

The skeleton of a hagfish is composed of cartilage, which includes a cartilaginous notochord, which runs the length of the body, and a skull. This notochord provides support to the fish’s body. Although they are craniates, hagfishes are not vertebrates, since they do not replace the notochord with a vertebral column during development, as do the vertebrates.
The clade Petromyzontidae includes approximately 40 species of lampreys. Lampreys are similar to hagfishes in size and shape; however, lampreys have a brain case and incomplete vertebrae. Lampreys lack paired appendages and bone, as do the hagfishes. As adults, lampreys are characterized by a toothed, funnel-like sucking mouth. Some species are parasitic as adults, attaching to and feeding on the body fluids of fish (Figure 2b). Most species are free-living.
Lampreys live primarily in coastal and fresh waters and have a worldwide temperate region distribution. All species spawn in fresh waters. Eggs are fertilized externally, and the larvae are distinctly different from the adult form, spending 3 to 15 years as suspension feeders. Once they attain sexual maturity, the adults reproduce and die within days. Lampreys have a notochord as adults.
Jawed Fishes
Gnathostomes or “jaw-mouths” are vertebrates that have jaws and include both cartilaginous and bony fishes. One of the most significant developments in early vertebrate evolution was the origin of the jaw, which is a hinged structure attached to the cranium that allows an animal to grasp and tear its food. The evolution of jaws allowed early gnathostomes to exploit food resources that were unavailable to jawless fishes.
The clade Chondrichthyes, the cartilaginous fishes, is diverse, consisting of sharks (Figure 3a), rays, and skates, together with sawfishes and a few dozen species of fishes called chimaeras, or ghost sharks. Chondrichthyes have paired fins and a skeleton made of cartilage. This clade arose approximately 370 million years ago in the middle Devonian. They are thought to have descended from an extinct group that had a skeleton made of bone; thus, the cartilaginous skeleton of Chondrichthyes is a later development. Parts of the shark skeleton are strengthened by granules of calcium carbonate, but this is not the same as bone.
Most cartilaginous fishes live in marine habitats, with a few species living in fresh water for some or all of their lives. Most sharks are carnivores that feed on live prey, either swallowing it whole or using their jaws and teeth to tear it into smaller pieces. Shark teeth likely evolved from the jagged scales that cover their skin. Some species of sharks and rays are suspension feeders that feed on plankton.

Sharks have well-developed sense organs that aid them in locating prey, including a keen sense of smell and electroreception, the latter being perhaps the most sensitive of any animal. Organs called ampullae of Lorenzini allow sharks to detect the electromagnetic fields that are produced by all living things, including their prey. Electroreception has only been observed in aquatic or amphibious animals. Sharks, together with most fishes, also have a sense organ called the lateral line, which is used to detect movement and vibration in the surrounding water, and a sense that is often considered homologous to “hearing” in terrestrial vertebrates. The lateral line is visible as a darker stripe that runs along the length of the fish’s body.
Sharks reproduce sexually and eggs are fertilized internally. Most species are ovoviviparous, that is, the fertilized egg is retained in the oviduct of the mother’s body, and the embryo is nourished by the egg yolk. The eggs hatch in the uterus and young are born alive and fully functional. Some species of sharks are oviparous: They lay eggs that hatch outside of the mother’s body. Embryos are protected by a shark egg case or “mermaid’s purse” that has the consistency of leather. The shark egg case has tentacles that snag in seaweed and give the newborn shark cover. A few species of sharks are viviparous, that is, the young develop within the mother’s body, and she gives live birth.
Rays and skates include more than 500 species and are closely related to sharks. They can be distinguished from sharks by their flattened bodies, pectoral fins that are enlarged and fused to the head, and gill slits on their ventral surface (Figure 3b). Like sharks, rays and skates have a cartilaginous skeleton. Most species are marine and live on the sea floor, with nearly a worldwide distribution.
Bony Fishes
Members of the clade Osteichthyes, or bony fishes, are characterized by a bony skeleton. The vast majority of present-day fishes belong to this group, which consists of approximately 30,000 species, making it the largest class of vertebrates in existence today.
Nearly all bony fishes have an ossified skeleton with specialized bone cells (osteocytes) that produce and maintain a calcium phosphate matrix. This characteristic has only reverted in a few groups of Osteichthyes, such as sturgeons and paddlefish, which have primarily cartilaginous skeletons. The skin of bony fishes is often covered in overlapping scales, and glands in the skin secrete mucus that reduces drag when swimming and aids the fish in osmoregulation. Like sharks, bony fishes have a lateral line system that detects vibrations in water. Unlike sharks, some bony fish depend on their eyesight to locate prey. Bony fish are also unusual in possessing taste cells in the head and trunk region of the body that allow them to detect extremely small concentrations of molecules in the water.
All bony fishes, like the cartilaginous fishes, use gills to breathe. Water is drawn over gills that are located in chambers covered and ventilated by a protective, muscular flap called the operculum. Unlike sharks, bony fishes have a swim bladder, a gas-filled organ that helps to control the buoyancy of the fish. Bony fishes are further divided into two clades with living members: Actinopterygii (ray-finned fishes) and Sarcopterygii (lobe-finned fishes).
The ray-finned fishes include many familiar fishes—tuna, bass, trout, and salmon (Figure 4a), among others. Ray-finned fishes are named for the form of their fins—webs of skin supported by bony spines called rays. In contrast, the fins of lobe-finned fishes are fleshy and supported by bone (Figure 4b). Living members of lobe-finned fishes include the less familiar lungfishes and coelacanth.

Amphibians
Amphibians are vertebrate tetrapods. Amphibia includes frogs, salamanders, and caecilians. The term amphibian means “dual life,” which is a reference to the metamorphosis that many frogs undergo from a tadpole to an adult and the mixture of aquatic and terrestrial environments in their life cycle. Amphibians evolved in the Devonian period and were the earliest terrestrial tetrapods.
As tetrapods, most amphibians are characterized by four well-developed limbs, although some species of salamanders and all caecilians possess only vestigial limbs. An important characteristic of extant amphibians is a moist, permeable skin, achieved by mucus glands. The moist skin allows oxygen and carbon dioxide exchange with the environment, a process called cutaneous respiration. All living adult amphibian species are carnivorous, and some terrestrial amphibians have a sticky tongue that is used to capture prey.
Amphibian Diversity
Amphibia comprise an estimated 6,500 extant species that inhabit tropical and temperate regions around the world. Amphibians can be divided into three clades: Urodela (“tailed-ones”), the salamanders and newts; Anura (“tail-less ones”), the frogs and toads; and Apoda (“legless ones”), the caecilians.
Living salamanders (Figure 5a) include approximately 500 species, some of which are aquatic, others terrestrial, and some that live on land only as adults. Adult salamanders usually have a generalized tetrapod body plan with four limbs and a tail. Some salamanders are lungless, and respiration occurs through the skin or external gills. Some terrestrial salamanders have primitive lungs; a few species have both gills and lungs.

Frogs (Figure 5b) are the most diverse group of amphibians, with approximately 5,000 species that live on all continents except Antarctica. Frogs have a body plan that is more specialized than the salamander body plan for movement on land. Adult frogs use their hind limbs to jump many times their body length on land. Frogs have a number of modifications that allow them to avoid predators, including skin that acts as camouflage and defensive chemicals that are poisonous to predators secreted from glands in the skin.
Frog eggs are fertilized externally, as they are laid in moist environments. Frogs demonstrate a range of parental behaviors, with some species exhibiting little care, to species that carry eggs and tadpoles on their hind legs or backs. The life cycle consists of two stages: the larval stage followed by metamorphosis to an adult stage. The larval stage of a frog, the tadpole, is often a filter-feeding herbivore. Tadpoles usually have gills, a lateral line system, long-finned tails, but no limbs. At the end of the tadpole stage, frogs undergo a gradual metamorphosis into the adult form. During this stage, the gills and lateral line system disappear, and four limbs develop. The jaws become larger and are suited for carnivorous feeding, and the digestive system transforms into the typical short gut of a predator. An eardrum and air-breathing lungs also develop. These changes during metamorphosis allow the larvae to move onto land in the adult stage (Figure 6).

Caecilians comprise an estimated 185 species. They lack external limbs and resemble giant earthworms. They inhabit soil and are found primarily in the tropics of South America, Africa, and southern Asia where they are adapted for a soil-burrowing lifestyle and are nearly blind. Unlike most of the other amphibians that breed in or near water, reproduction in a drier soil habitat means that caecilians must utilize internal fertilization, and most species give birth to live young (Figure 7).

Reptiles and Birds
The amniotes—reptiles, birds, and mammals—are distinguished from amphibians by their terrestrially adapted (shelled) egg and an embryo protected by amniotic membranes. The evolution of amniotic membranes meant that the embryos of amniotes could develop within an aquatic environment inside the egg. This led to less dependence on a water environment for development and allowed the amniotes to invade drier areas. This was a significant evolutionary change that distinguished them from amphibians, which were restricted to moist environments due to their shell-less eggs. Although the shells of various amniotic species vary significantly, they all allow retention of water. The membranes of the amniotic egg also allowed gas exchange and sequestering of wastes within the enclosure of an eggshell. The shells of bird eggs are composed of calcium carbonate and are hard and brittle, but possess pores for gas and water exchange. The shells of reptile eggs are more leathery and pliable. Most mammals do not lay eggs; however, even with internal gestation, amniotic membranes are still present.
In the past, the most common division of amniotes has been into classes Mammalia, Reptilia, and Aves. Birds are descended, however, from dinosaurs, so this classical scheme results in groups that are not true clades. We will discuss birds as a group distinct from reptiles with the understanding that this does not reflect evolutionary history.
Reptiles
Reptiles are tetrapods. Limbless reptiles—snakes—may have vestigial limbs and, like caecilians, are classified as tetrapods because they are descended from four-limbed ancestors. Reptiles lay shelled eggs on land. Even aquatic reptiles, like sea turtles, return to the land to lay eggs. They usually reproduce sexually with internal fertilization. Some species display ovoviviparity, with the eggs remaining in the mother’s body until they are ready to hatch. Other species are viviparous, with the offspring born alive.
One of the key adaptations that permitted reptiles to live on land was the development of their scaly skin, containing the protein keratin and waxy lipids, which prevented water loss from the skin. This occlusive skin means that reptiles cannot use their skin for respiration, like amphibians, and thus all must breathe with lungs. In addition, reptiles conserve valuable body water by excreting nitrogen in the form of uric acid paste. These characteristics, along with the shelled, amniotic egg, were the major reasons why reptiles became so successful in colonizing a variety of terrestrial habitats far from water.
Reptiles are ectotherms, that is, animals whose main source of body heat comes from the environment. Behavioral maneuvers, like basking to heat themselves, or seeking shade or burrows to cool off, help them regulate their body temperature,
Class Reptilia includes diverse species classified into four living clades. These are the Crocodilia, Sphenodontia, Squamata, and Testudines.
The Crocodilia (“small lizard”) arose approximately 84 million years ago, and living species include alligators, crocodiles, and caimans. Crocodilians (Figure 8a) live throughout the tropics of Africa, South America, the southeastern United States, Asia, and Australia. They are found in freshwater habitats, such as rivers and lakes, and spend most of their time in water. Some species are able to move on land due to their semi-erect posture.

The Sphenodontia (“wedge tooth”) arose in the Mesozoic Era and includes only one living genus, Tuatara, with two species that are found in New Zealand. There are many fossil species extending back to the Triassic period (250–200 million years ago). Although the tuataras resemble lizards, they are anatomically distinct and share characteristics that are found in birds and turtles.
Squamata (“scaly”) arose in the late Permian; living species include lizards and snakes, which are the largest extant clade of reptiles (Figure 8b). Lizards differ from snakes by having four limbs, eyelids, and external ears, which are lacking in snakes. Lizard species range in size from chameleons and geckos that are a few centimeters in length to the Komodo dragon, which is about 3 meters in length.
Snakes are thought to have descended from either burrowing lizards or aquatic lizards over 100 million years ago (Figure 8c). Snakes comprise about 3,000 species and are found on every continent except Antarctica. They range in size from 10 centimeter-long thread snakes to 7.5 meter-long pythons and anacondas. All snakes are carnivorous and eat small animals, birds, eggs, fish, and insects.
Turtles are members of the clade Testudines (“having a shell”) (Figure 8d). Turtles are characterized by a bony or cartilaginous shell, made up of the carapace on the back and the plastron on the ventral surface, which develops from the ribs. Turtles arose approximately 200 million years ago, predating crocodiles, lizards, and snakes. Turtles lay eggs on land, although many species live in or near water. Turtles range in size from the speckled padloper tortoise at 8 centimeters (3.1 inches) to the leatherback sea turtle at 200 centimeters (over 6 feet). The term “turtle” is sometimes used to describe only those species of Testudines that live in the sea, with the terms “tortoise” and “terrapin” used to refer to species that live on land and in fresh water, respectively.
Birds
Data now suggest that birds belong within the reptile clade, but they display a number of unique adaptations that set them apart. Unlike the reptiles, birds are endothermic, meaning they generate their own body heat through metabolic processes. The most distinctive characteristic of birds is their feathers, which are modified reptilian scales. Birds have several different types of feathers that are specialized for specific functions, like contour feathers that streamline the bird’s exterior and loosely structured down feathers that insulate (Figure 9a).
Feathers not only permitted the earliest birds to glide, and ultimately engage in flapping flight, but they insulated the bird’s body, assisting the maintenance of endothermy, even in cooler temperatures. Powering a flying animal requires economizing on the amount of weight carried. As body weight increases, the muscle output and energetic cost required for flying increase. Birds have made several modifications to reduce body weight, including hollow or pneumatic bones (Figure 9b) with air spaces that may be connected to air sacs and cross-linked struts within their bones to provide structural reinforcement. Parts of the vertebral skeleton and braincase are fused to increase its strength while lightening its weight. Most species of bird only possess one ovary rather than two, and no living birds have teeth in their jaw, further reducing body mass.
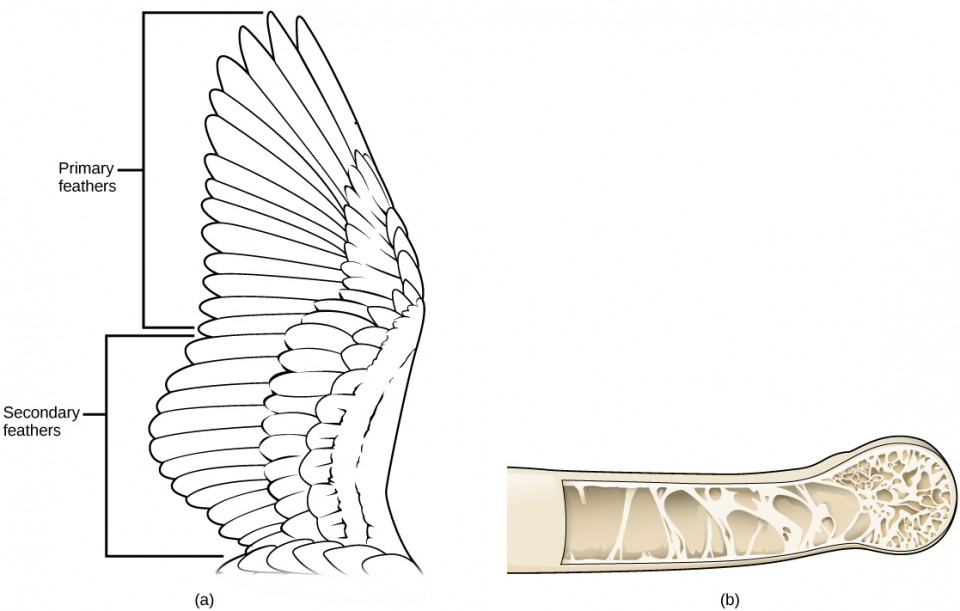
Birds possess a system of air sacs branching from their primary airway that divert the path of air so that it passes unidirectionally through the lung, during both inspiration and expiration. Unlike mammalian lungs in which air flows in two directions as it is breathed in and out, air flows continuously through the bird’s lung to provide a more efficient system of gas exchange.
Mammals
Mammals are vertebrates that have hair and mammary glands used to provide nutrition for their young. Certain features of the jaw, skeleton, skin, and internal anatomy are also unique to mammals. The presence of hair is one of the key characteristics of a mammal. Although it is not very extensive in some groups, such as whales, hair has many important functions for mammals. Mammals are endothermic, and hair provides insulation by trapping a layer of air close to the body to retain metabolic heat. Hair also serves as a sensory mechanism through specialized hairs called vibrissae, better known as whiskers. These attach to nerves that transmit touch information, which is particularly useful to nocturnal or burrowing mammals. Hair can also provide protective coloration.
Mammalian skin includes secretory glands with various functions. Sebaceous glands produce a lipid mixture called sebum that is secreted onto the hair and skin for water resistance and lubrication. Sebaceous glands are located over most of the body. Sudoriferous glands produce sweat and scent, which function in thermoregulation and communication, respectively. Mammary glands produce milk that is used to feed newborns. While male monotremes and eutherians possess mammary glands, male marsupials do not.
The skeletal system of mammals possesses unique features that differentiate them from other vertebrates. Most mammals have heterodont teeth, meaning they have different types and shapes of teeth that allow them to feed on different kinds of foods. These different types of teeth include the incisors, the canines, premolars, and molars. The first two types are for cutting and tearing, whereas the latter two types are for crushing and grinding. Different groups have different proportions of each type, depending on their diet. Most mammals are also diphyodonts, meaning they have two sets of teeth in their lifetime: deciduous or “baby” teeth, and permanent teeth. In other vertebrates, the teeth can be replaced throughout life.
Modern mammals are divided into three broad groups: monotremes, marsupials, and eutherians (or placental mammals). The eutherians, or placental mammals, and the marsupials collectively are called therian mammals, whereas monotremes are called metatherians.
There are three living species of monotremes: the platypus and two species of echidnas, or spiny anteaters (Figure 10). The platypus and one species of echidna are found in Australia, whereas the other species of echidna is found in New Guinea. Monotremes are unique among mammals, as they lay leathery eggs, similar to those of reptiles, rather than giving birth to live young. However, the eggs are retained within the mother’s reproductive tract until they are almost ready to hatch. Once the young hatch, the female begins to secrete milk from pores in a ridge of mammary tissue along the ventral side of her body. Like other mammals, monotremes are endothermic but regulate body temperatures somewhat lower (90 °F, 32 °C) than placental mammals do (98 °F, 37 °C). Like reptiles, monotremes have one posterior opening for urinary, fecal, and reproductive products, rather than three separate openings like placental mammals do. Adult monotremes lack teeth.

Marsupials are found primarily in Australia and nearby islands, although about 100 species of opossums and a few species of two other families are found in the Americas. Australian marsupials number over 230 species and include the kangaroo, koala, bandicoot, and Tasmanian devil (Figure 11). Most species of marsupials possess a pouch in which the young reside after birth, receiving milk and continuing to develop. Before birth, marsupials have a less complex placental connection, and the young are born much less developed than in placental mammals.

Eutherians are the most widespread of the mammals, occurring throughout the world. There are several groups of eutherians, including Insectivora, the insect eaters; Edentata, the toothless anteaters; Rodentia, the rodents; Chiroptera, the bats; Cetacea, the aquatic mammals including whales; Carnivora, carnivorous mammals including dogs, cats, and bears; and Primates, which includes humans. Eutherian mammals are sometimes called placental mammals, because all species have a complex placenta that connects a fetus to the mother, allowing for gas, fluid, waste, and nutrient exchange. While other mammals may possess a less complex placenta or briefly have a placenta, all eutherians have a complex placenta during gestation.
Primates
Order Primates of class Mammalia includes lemurs, tarsiers, monkeys, and the apes, which include humans. Non-human primates live primarily in tropical or subtropical regions of South America, Africa, and Asia. They range in size from the mouse lemur at 30 grams (1 ounce) to the mountain gorilla at 200 kilograms (441 pounds). The characteristics and evolution of primates are of particular interest to us as they allow us to understand the evolution of our own species.
All primate species have adaptations for climbing trees, as they all descended from tree-dwellers, although not all species are arboreal. This arboreal heritage of primates resulted in hands and feet that are adapted for brachiation, or climbing and swinging through trees. These adaptations include, but are not limited to 1) a rotating shoulder joint, 2) a big toe that is widely separated from the other toes and thumbs that are widely separated from fingers (except humans), which allow for gripping branches, and 3) stereoscopic vision, two overlapping visual fields, which allows for the depth perception necessary to gauge distance. Other characteristics of primates are brains that are larger than those of many other mammals, claws that have been modified into flattened nails, typically only one offspring per pregnancy, and a trend toward holding the body upright.
Order Primates is divided into two groups: prosimians and anthropoids. Prosimians include the bush babies of Africa, the lemurs of Madagascar, and the lorises, pottos, and tarsiers of Southeast Asia. Anthropoids include monkeys, lesser apes, and great apes (Figure 12). In general, prosimians tend to be nocturnal, smaller in size than anthropoids, and have relatively smaller brains compared to anthropoids.

- Actinopterygii
- ray-finned fishes
- amniote
- a clade of animals that possesses an amniotic egg; includes reptiles (including birds) and mammals
- Amphibia
- frogs, salamanders, and caecilians
- ampulla of Lorenzini
- a sensory organ that allows sharks to detect electromagnetic fields produced by living things
- anthropoids
- a clade consisting of monkeys, apes, and humans
- Anura
- frogs
- Apoda
- caecilians
- brachiation
- swinging through trees
- caecilian
- a legless amphibian that belongs to clade Apoda
- Chondrichthyes
- jawed fishes with paired fins and a skeleton made of cartilage
- craniate
- a proposed clade of chordates that includes all groups except the tunicates and lancelets
- Crocodilia
- crocodiles and alligators
- cutaneous respiration
- gas exchange through the skin
- diphyodont
- refers to the possession of two sets of teeth in a lifetime
- down feather
- feather specialized for insulation
- eutherian mammal
- a mammal with a complex placenta, which connects a fetus to the mother; sometimes called placental mammals
- frog
- a tail-less amphibian that belongs to clade Anura
- gnathostome
- a jawed fish
- hagfish
- an eel-like jawless fish that lives on the ocean floor and is a scavenger
- heterodont teeth
- different types of teeth modified by different purposes
- lamprey
- a jawless fish characterized by a toothed, funnel-like, sucking mouth
- lateral line
- the sense organ that runs the length of a fish’s body, used to detect vibration in the water
- mammal
- one of the groups of endothermic vertebrates that possess hair and mammary glands
- mammary gland
- in female mammals, a gland that produces milk for newborns
- marsupial
- one of the groups of mammals that includes the kangaroo, koala, bandicoot, Tasmanian devil, and several other species; young develop within a pouch
- monotreme
- an egg-laying mammal
- Myxini
- hagfishes
- Osteichthyes
- bony fishes
- ostracoderm
- one of the earliest jawless fishes covered in bone
- Petromyzontidae
- the clade of lampreys
- pneumatic bone
- an air-filled bone
- Primates
- includes lemurs, tarsiers, monkeys, apes, and humans
- prosimians
- a group of primates that includes bush babies of Africa, lemurs of Madagascar, and lorises, pottos, and tarsiers of southeast Asia
- salamander
- a tailed amphibian that belongs to the clade Urodela
- Sarcopterygii
- lobe-finned fishes
- sebaceous gland
- in mammals, a skin gland that produce a lipid mixture called sebum
- Sphenodontia
- the reptilian clade that includes the tuataras
- Squamata
- the reptilian clade of lizards and snakes
- stereoscopic vision
- two overlapping fields of vision from the eyes that produces depth perception
- sudoriferous gland
- a gland in mammals that produces sweat and scent molecules
- swim bladder
- in fishes, a gas filled organ that helps to control the buoyancy of the fish
- tadpole
- the larval stage of a frog
- Testudines
- turtles
- Urodela
- salamanders
Glossary
Access for free at https://openstax.org/books/concepts-biology/pages/1-introduction
