28 Population Genetics
By the end of this section, you will be able to do the following:
- Describe the different types of variation in a population
- Explain why only natural selection can act upon heritable variation
- Describe genetic drift and the bottleneck effect
- Explain how each evolutionary force can influence a population’s allele frequencies
A population’s individuals often display different phenotypes, or express different alleles of a particular gene, which scientists refer to as polymorphisms. We call populations with two or more variations of particular characteristics polymorphic. A number of factors, including the population’s genetic structure and the environment (Figure 1) influence population variation, the distribution of phenotypes among individuals. Understanding phenotypic variation sources in a population is important for determining how a population will evolve in response to different evolutionary pressures.

Genetic Variance
Natural selection and some of the other evolutionary forces can only act on heritable traits, namely an organism’s genetic code. Because alleles are passed from parent to offspring, those that confer beneficial traits or behaviors may be selected, while deleterious alleles may not. Acquired traits, for the most part, are not heritable. For example, if an athlete works out in the gym every day, building up muscle strength, the athlete’s offspring will not necessarily grow up to be a body builder. If there is a genetic basis for the ability to run fast, on the other hand, a parent may pass this to a child.
LINK TO LEARNING
Before Darwinian evolution became the prevailing theory of the field, French naturalist Jean-Baptiste Lamarck theorized that organisms could inherit acquired traits. While the majority of scientists have not supported this hypothesis, some have recently begun to realize that Lamarck was not completely wrong. Read this article to learn more.
Heritability is the fraction of phenotype variation that we can attribute to genetic differences, or genetic variance, among individuals in a population. The greater the heritability of a population’s phenotypic variation, the more susceptible it is to the evolutionary forces that act on heritable variation.
We call the diversity of alleles and genotypes within a population genetic variance. When scientists are involved in the breeding of a species, such as with animals in zoos and nature preserves, they try to increase a population’s genetic variance to preserve as much of the phenotypic diversity as possible. This also helps reduce associated risks of inbreeding, the mating of closely related individuals, which can have the undesirable effect of bringing together deleterious recessive mutations that can cause abnormalities and susceptibility to disease. For example, a disease that is caused by a rare, recessive allele might exist in a population, but it will only manifest itself when an individual carries two copies of the allele. Because the allele is rare in a normal, healthy population with unrestricted habitat, the chance that two carriers will mate is low, and even then, only 25 percent of their offspring will inherit the disease allele from both parents. While it is likely to happen at some point, it will not happen frequently enough for natural selection to be able to swiftly eliminate the allele from the population, and as a result, the allele maintains itself at low levels in the gene pool. However, if a family of carriers begins to interbreed with each other, this will dramatically increase the likelihood of two carriers mating and eventually producing diseased offspring, a phenomenon that scientists call inbreeding depression.
Changes in allele frequencies that we identify in a population can shed light on how it is evolving. In addition to natural selection, there are other evolutionary forces that could be in play: genetic drift, gene flow, mutation, nonrandom mating, and environmental variances.
Genetic Drift
The theory of natural selection stems from the observation that some individuals in a population are more likely to survive longer and have more offspring than others; thus, they will pass on more of their genes to the next generation. A big, powerful male gorilla, for example, is much more likely than a smaller, weaker one to become the population’s silverback, the pack’s leader who mates far more than the other males of the group. The pack leader will father more offspring, who share half of his genes, and are likely to also grow bigger and stronger like their father. Over time, the genes for bigger size will increase in frequency in the population, and the population will, as a result, grow larger on average. That is, this would occur if this particular selection pressure, or driving selective force, were the only one acting on the population. In other examples, better camouflage or a stronger resistance to drought might pose a selection pressure.
Another way a population’s allele and genotype frequencies can change is genetic drift (Figure 2), which is simply the effect of chance. By chance, some individuals will have more offspring than others—not due to an advantage conferred by some genetically-encoded trait, but just because one male happened to be in the right place at the right time (when the receptive female walked by) or because the other one happened to be in the wrong place at the wrong time (when a fox was hunting).
VISUAL CONNECTION

Do you think genetic drift would happen more quickly on an island or on the mainland?
Answer:
Genetic drift is likely to occur more rapidly on an island where smaller populations are expected to occur.
Small populations are more susceptible to the forces of genetic drift. Large populations, alternatively, are buffered against the effects of chance. If one individual of a population of 10 individuals happens to die at a young age before it leaves any offspring to the next generation, all of its genes—1/10 of the population’s gene pool—will be suddenly lost. In a population of 100, that’s only 1 percent of the overall gene pool; therefore, it is much less impactful on the population’s genetic structure.
LINK TO LEARNING
Watch an animation of random sampling and genetic drift in action.
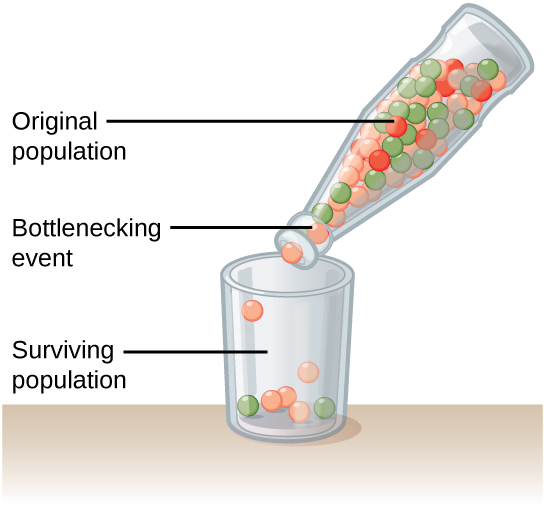
Natural events, such as an earthquake disaster that kills—at random—a large portion of the population, can magnify genetic drift. Known as the bottleneck effect, it results in suddenly wiping out a large portion of the genome (Figure 3). At once, the survivors’ genetic structure becomes the entire population’s genetic structure, which may be very different from the pre-disaster population.
Another scenario in which populations might experience a strong influence of genetic drift is if some portion of the population leaves to start a new population in a new location or if a physical barrier divides a population. In this situation, those individuals are an unlikely representation of the entire population, which results in the founder effect. The founder effect occurs when the genetic structure changes to match that of the new population’s founding fathers and mothers. Researchers believe that the founder effect was a key factor in the genetic history of the Afrikaner population of Dutch settlers in South Africa, as evidenced by mutations that are common in Afrikaners but rare in most other populations. This is probably because a higher-than-normal proportion of the founding colonists carried these mutations. As a result, the population expresses unusually high incidences of Huntington’s disease (HD) and Fanconi anemia (FA), a genetic disorder known to cause blood marrow and congenital abnormalities—even cancer.1
SCIENTIFIC METHOD CONNECTION
Testing the Bottleneck Effect
Question: How do natural disasters affect a population’s genetic structure?
Background: When an earthquake or hurricane suddenly wipes out much of a population, the surviving individuals are usually a random sampling of the original group. As a result, the population’s genetic makeup can change dramatically. We call this phenomenon the bottleneck effect.
Hypothesis: Repeated natural disasters will yield different population genetic structures; therefore, each time one runs this experiment the results will vary.
Test the hypothesis: Count out the original population using different colored beads. For example, red, blue, and yellow beads might represent red, blue, and yellow individuals. After recording the number of each individual in the original population, place them all in a bottle with a narrow neck that will only allow a few beads out at a time. Then, pour 1/3 of the bottle’s contents into a bowl. This represents the surviving individuals after a natural disaster kills a majority of the population. Count the number of the different colored beads in the bowl, and record it. Then, place all of the beads back in the bottle and repeat the experiment four more times.
Analyze the data: Compare the five populations that resulted from the experiment. Do the populations all contain the same number of different colored beads, or do they vary? Remember, these populations all came from the same exact parent population.
Form a conclusion: Most likely, the five resulting populations will differ quite dramatically. This is because natural disasters are not selective—they kill and spare individuals at random. Now think about how this might affect a real population. What happens when a hurricane hits the Mississippi Gulf Coast? How do the seabirds that live on the beach fare?
Gene Flow
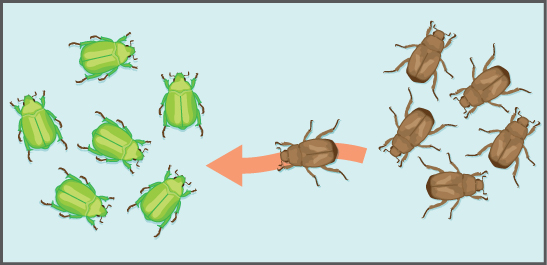
Another important evolutionary force is gene flow: the flow of alleles in and out of a population due to the migration of individuals or gametes (Figure 4). While some populations are fairly stable, others experience more flux. Many plants, for example, send their pollen far and wide, by wind or by bird, to pollinate other populations of the same species some distance away. Even a population that may initially appear to be stable, such as a pride of lions, can experience its fair share of immigration and emigration as developing males leave their mothers to seek out a new pride with genetically unrelated females. This variable flow of individuals in and out of the group not only changes the population’s gene structure, but it can also introduce new genetic variation to populations in different geological locations and habitats.
Mutation
Mutations are changes to an organism’s DNA and are an important driver of diversity in populations. Species evolve because of mutations accumulating over time. The appearance of new mutations is the most common way to introduce novel genotypic and phenotypic variance. Some mutations are unfavorable or harmful and are quickly eliminated from the population by natural selection. Others are beneficial and will spread through the population. Whether or not a mutation is beneficial or harmful is determined by whether it helps an organism survive to sexual maturity and reproduce. Some mutations do not do anything and can linger, unaffected by natural selection, in the genome. Some can have a dramatic effect on a gene and the resulting phenotype.
Nonrandom Mating
If individuals nonrandomly mate with their peers, the result can be a changing population. There are many reasons nonrandom mating occurs. One reason is simple mate choice. For example, female peahens may prefer peacocks with bigger, brighter tails. Natural selection picks traits that lead to more mating selections for an individual. One common form of mate choice, called assortative mating, is an individual’s preference to mate with partners who are phenotypically similar to themselves.
Another cause of nonrandom mating is physical location. This is especially true in large populations spread over vast geographic distances where not all individuals will have equal access to one another. Some might be miles apart through woods or over rough terrain, while others might live immediately nearby.
Environmental Variance
Genes are not the only players involved in determining population variation. Other factors, such as the environment (Figure 5) also influence phenotypes. A beachgoer is likely to have darker skin than a city dweller, for example, due to regular exposure to the sun, an environmental factor. For some species, the environment determines some major characteristics, such as gender. For example, some turtles and other reptiles have temperature-dependent sex determination (TSD). TSD means that individuals develop into males if their eggs are incubated within a certain temperature range, or females at a different temperature range.

Geographic separation between populations can lead to differences in the phenotypic variation between those populations. We see such geographical variation between most populations and it can be significant. We can observe one type of geographic variation, a cline, as given species’ populations vary gradually across an ecological gradient. Species of warm-blooded animals, for example, tend to have larger bodies in the cooler climates closer to the earth’s poles, allowing them to better conserve heat. This is a latitudinal cline. Alternatively, flowering plants tend to bloom at different times depending on where they are along a mountain slope. This is an altitudinal cline.
If there is gene flow between the populations, the individuals will likely show gradual differences in phenotype along the cline. Restricted gene flow, alternatively can lead to abrupt differences, even speciation.
Footnotes
- 1 A. J. Tipping et al., “Molecular and Genealogical Evidence for a Founder Effect in Fanconi Anemia Families of the Afrikaner Population of South Africa,” PNAS 98, no. 10 (2001): 5734-5739, doi: 10.1073/pnas.091402398.
Glossary
- assortative mating
- when individuals tend to mate with those who are phenotypically similar to themselves
- bottleneck effect
- magnification of genetic drift as a result of natural events or catastrophes
- cline
- gradual geographic variation across an ecological gradient
- gene flow
- flow of alleles in and out of a population due to the individual or gamete migration
- genetic drift
- effect of chance on a population’s gene pool
- genetic variance
- diversity of alleles and genotypes in a population
- geographical variation
- differences in the phenotypic variation between populations that are separated geographically
- heritability
- fraction of population variation that can be attributed to its genetic variance
- inbreeding
- mating of closely related individuals
- inbreeding depression
- increase in abnormalities and disease in inbreeding populations
- nonrandom mating
- changes in a population’s gene pool due to mate choice or other forces that cause individuals to mate with certain phenotypes more than others
- population variation
- distribution of phenotypes in a population
- selective pressure
- environmental factor that causes one phenotype to be better than another
Access for free at https://openstax.org/books/biology-2e/pages/1-introduction

{kind=link}