34 Perspectives on the Phylogenetic Tree
By the end of this section, you will be able to do the following:
- Describe horizontal gene transfer
- Illustrate how prokaryotes and eukaryotes transfer genes horizontally
- Identify the web and ring models of phylogenetic relationships and describe how they differ from the original phylogenetic tree concept
Phylogenetic modeling concepts are constantly changing. It is one of the most dynamic fields of study in all biology. Over the last several decades, new research has challenged scientists’ ideas about how organisms are related. The scientific community has proposed new models of these relationships.
Many phylogenetic trees are models of the evolutionary relationship among species. Phylogenetic trees originated with Charles Darwin, who sketched the first phylogenetic tree in 1837 (Figure 1a). This served as a prototype for subsequent studies for more than a century. The phylogenetic tree concept with a single trunk representing a common ancestor, with the branches representing the divergence of species from this ancestor, fits well with the structure of many common trees, such as the oak (Figure 1b). However, evidence from modern DNA sequence analysis and newly developed computer algorithms has caused skepticism about the standard tree model’s validity in the scientific community.

Limitations to the Classic Model
Classical thinking about prokaryotic evolution, included in the classic tree model, is that species evolve clonally. That is, they produce offspring themselves with only random mutations causing the descent into the variety of modern-day and extinct species known to science. This view is somewhat complicated in eukaryotes that reproduce sexually, but the laws of Mendelian genetics explain the variation in offspring, again, to be a result of a mutation within the species. Scientists did not consider the concept of genes transferring between unrelated species as a possibility until relatively recently. Horizontal gene transfer (HGT), or lateral gene transfer, is the transfer of genes between unrelated species. HGT is an ever-present phenomenon, with many evolutionists postulating a major role for this process in evolution, thus complicating the simple tree model. Genes pass between species which are only distantly related using standard phylogeny, thus adding a layer of complexity to understanding phylogenetic relationships.
The various ways that HGT occurs in prokaryotes is important to understanding phylogenies. Although at present some do not view HGT as important to eukaryotic evolution, HGT does occur in this domain as well. Finally, as an example of the ultimate gene transfer, some scientists have proposed genome fusion theories between symbiotic or endosymbiotic organisms to explain an event of great importance—the evolution of the first eukaryotic cell, without which humans could not have come into existence.
Horizontal Gene Transfer
Horizontal gene transfer (HGT) is the introduction of genetic material from one species to another species by mechanisms other than the vertical transmission from parent(s) to offspring. These transfers allow even distantly related species to share genes, influencing their phenotypes. Scientists believe that HGT is more prevalent in prokaryotes, but that this process transfers only about 2% of the prokaryotic genome. Some researchers believe such estimates are premature: we must view the actual importance of HGT to evolutionary processes as a work in progress. As scientists investigate this phenomenon more thoroughly, they may reveal more HGT transfer. Many scientists believe that HGT and mutation are (especially in prokaryotes) a significant source of genetic variation, which is the raw material in the natural selection process. These transfers may occur between any two species that share an intimate relationship (Table 1).
| Mechanism | Mode of Transmission | Example | |
|---|---|---|---|
| Prokaryotes | transformation | DNA uptake | many prokaryotes |
| transduction | bacteriophage (virus) | bacteria | |
| conjugation | pilus | many prokaryotes | |
| gene transfer agents | phage-like particles | purple non-sulfur bacteria | |
| Eukaryotes | from food organisms | unknown | aphid |
| jumping genes | transposons | rice and millet plants | |
| epiphytes/parasites | unknown | yew tree fungi | |
| from viral infections |
HGT in Prokaryotes
HGT mechanisms are quite common in the Bacteria and Archaea domains, thus significantly changing the way scientists view their evolution. The majority of evolutionary models, such as in the Endosymbiont Theory, propose that eukaryotes descended from multiple prokaryotes, which makes HGT all the more important to understanding the phylogenetic relationships of all extant and extinct species. The Endosymbiont Theory purports that the eukaryotes’ mitochondria and the green plants’ chloroplasts and flagellates originated as free-living prokaryotes that invaded primitive eukaryotic cells and become established as permanent symbionts in the cytoplasm.
Microbiology students are well aware that genes transfer among common bacteria. These gene transfers between species are the major mechanism whereby bacteria acquire resistance to antibiotics. Classically, scientists believe that three different mechanisms drive such transfers.
- Transformation: bacteria takes up naked DNA
- Transduction: a virus transfers the genes
- Conjugation: a hollow tube, or pilus transfers genes between organisms
More recently, scientists have discovered a fourth gene transfer mechanism between prokaryotes. Small, virus-like particles, or gene transfer agents (GTAs) transfer random genomic segments from one prokaryote species to another. GTAs are responsible for genetic changes, sometimes at a very high frequency compared to other evolutionary processes. Scientists characterized the first GTA in 1974 using purple, non-sulfur bacteria. These GTAs, which are most likely bacteriophages that lost the ability to reproduce on their own, carry random DNA pieces from one organism to another. Controlled studies using marine bacteria have demonstrated GTAs’ ability to act with high frequency. Scientists have estimated gene transfer events in marine prokaryotes, either by GTAs or by viruses, to be as high as 1013 per year in the Mediterranean Sea alone. GTAs and viruses are efficient HGT vehicles with a major impact on prokaryotic evolution.
As a consequence of this modern DNA analysis, the idea that eukaryotes evolved directly from Archaea has fallen out of favor. While eukaryotes share many features that are absent in bacteria, such as the TATA box (located in many genes’ promoter region), the discovery that some eukaryotic genes were more homologous with bacterial DNA than Archaea DNA made this idea less tenable. Furthermore, scientists have proposed genome fusion from Archaea and Bacteria by endosymbiosis as the ultimate event in eukaryotic evolution.
HGT in Eukaryotes
Although it is easy to see how prokaryotes exchange genetic material by HGT, scientists initially thought that this process was absent in eukaryotes. After all, prokaryotes are but single cells exposed directly to their environment; whereas, the multicellular organisms’ sex cells are usually sequestered in protected parts of the body. It follows from this idea that the gene transfers between multicellular eukaryotes should be more difficult. Scientists believe this process is rarer in eukaryotes and has a much smaller evolutionary impact than in prokaryotes. In spite of this, HGT between distantly related organisms is evident in several eukaryotic species, and it is possible that scientists will discover more examples in the future.
In plants, researchers have observed gene transfer in species that cannot cross-pollinate by normal means. Transposons or “jumping genes” have shown a transfer between rice and millet plant species. Furthermore, fungal species feeding on yew trees, from which the anti-cancer drug TAXOL® is derived from the bark, have acquired the ability to make taxol themselves, a clear example of gene transfer.
In animals, a particularly interesting example of HGT occurs within the aphid species (Figure 2). Aphids are insects that vary in color based on carotenoid content. Carotenoids are pigments that a variety of plants, fungi, and microbes produce, and they serve a variety of functions in animals, who obtain these chemicals from their food. Humans require carotenoids to synthesize vitamin A, and we obtain them by eating orange fruits and vegetables: carrots, apricots, mangoes, and sweet potatoes. Alternatively, aphids have acquired the ability to make the carotenoids on their own. According to DNA analysis, this ability is due to fungal genes transferring into the insect by HGT, presumably as the insect consumed fungi for food. A carotenoid enzyme, or desaturase, is responsible for the red coloration in certain aphids, and when mutation activates this gene, the aphids revert to their more common green color (Figure 2).

Genome Fusion and Eukaryote Evolution
Scientists believe the ultimate in HGT occurs through genome fusion between different prokaryote species when two symbiotic organisms become endosymbiotic. This occurs when one species is taken inside another species’ cytoplasm, which ultimately results in a genome consisting of genes from both the endosymbiont and the host. This mechanism is an aspect of the Endosymbiont Theory, which most biologists accept as the mechanism whereby eukaryotic cells obtained their mitochondria and chloroplasts. However, the role of endosymbiosis in developing the nucleus is more controversial. Scientists believe that nuclear and mitochondrial DNA have different (separate) evolutionary origins, with the mitochondrial DNA derived from the bacteria’s circular genomes ancient prokaryotic cells engulfed. We can regard mitochondrial DNA as the smallest chromosome. Interestingly enough, mitochondrial DNA is inherited only from the mother. The mitochondrial DNA degrades in sperm when the sperm degrades in the fertilized egg or in other instances when the mitochondria located in the sperm’s flagellum fails to enter the egg.
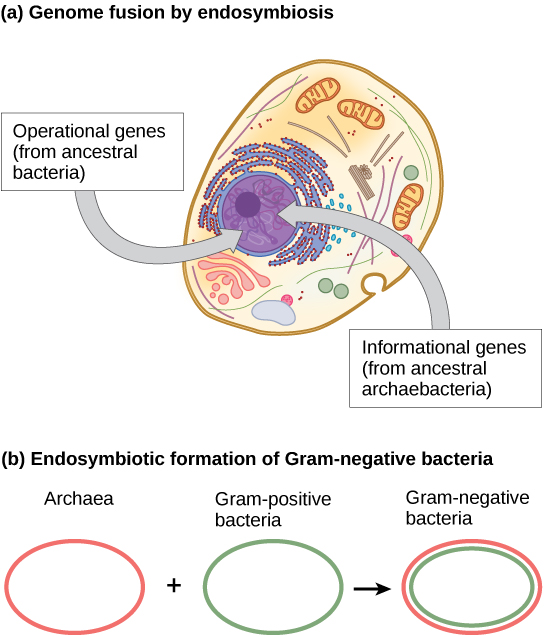
Within the past decade, James Lake of the UCLA/NASA Astrobiology Institute proposed that the genome fusion process is responsible for the evolution of the first eukaryotic cells (Figure 3a). Using DNA analysis and a new mathematical algorithm, conditioned reconstruction (CR), his laboratory proposed that eukaryotic cells developed from an endosymbiotic gene fusion between two species, one an Archaea and the other a Bacteria. As mentioned, some eukaryotic genes resemble those of Archaea; whereas, others resemble those from Bacteria. An endosymbiotic fusion event, such as Lake has proposed, would clearly explain this observation. Alternatively, this work is new and the CR algorithm is relatively unsubstantiated, which causes many scientists to resist this hypothesis.
Lake’s more recent work (Figure 3b) proposes that gram-negative bacteria, which are unique within their domain in that they contain two lipid bilayer membranes, resulted from an endosymbiotic fusion of archaeal and bacterial species. The double membrane would be a direct result of the endosymbiosis, with the endosymbiont picking up the second membrane from the host as it was internalized. Scientists have also used this mechanism to explain the double membranes in mitochondria and chloroplasts. Lake’s work is not without skepticism, and the biological science community still debates his ideas. In addition to Lake’s hypothesis, there are several other competing theories as to the origin of eukaryotes. How did the eukaryotic nucleus evolve? One theory is that the prokaryotic cells produced an additional membrane that surrounded the bacterial chromosome. Some bacteria have the DNA enclosed by two membranes; however, there is no evidence of a nucleolus or nuclear pores. Other proteobacteria also have membrane-bound chromosomes. If the eukaryotic nucleus evolved this way, we would expect one of the two types of prokaryotes to be more closely related to eukaryotes.
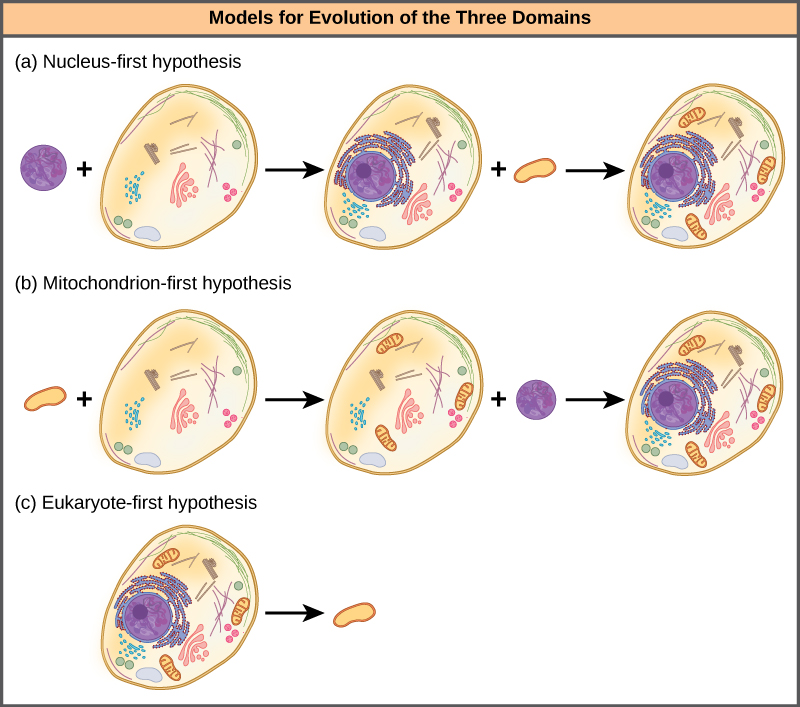
The nucleus-first hypothesis proposes that the nucleus evolved in prokaryotes first (Figure 4a), followed by a later fusion of the new eukaryote with bacteria that became mitochondria. The mitochondria-first hypothesis proposes that mitochondria were first established in a prokaryotic host (Figure 4b), which subsequently acquired a nucleus, by fusion or other mechanisms, to become the first eukaryotic cell. Most interestingly, the eukaryote-first hypothesis proposes that prokaryotes actually evolved from eukaryotes by losing genes and complexity (Figure 4c). All of these hypotheses are testable. Only time and more experimentation will determine which hypothesis data best supports.
Web and Network Models
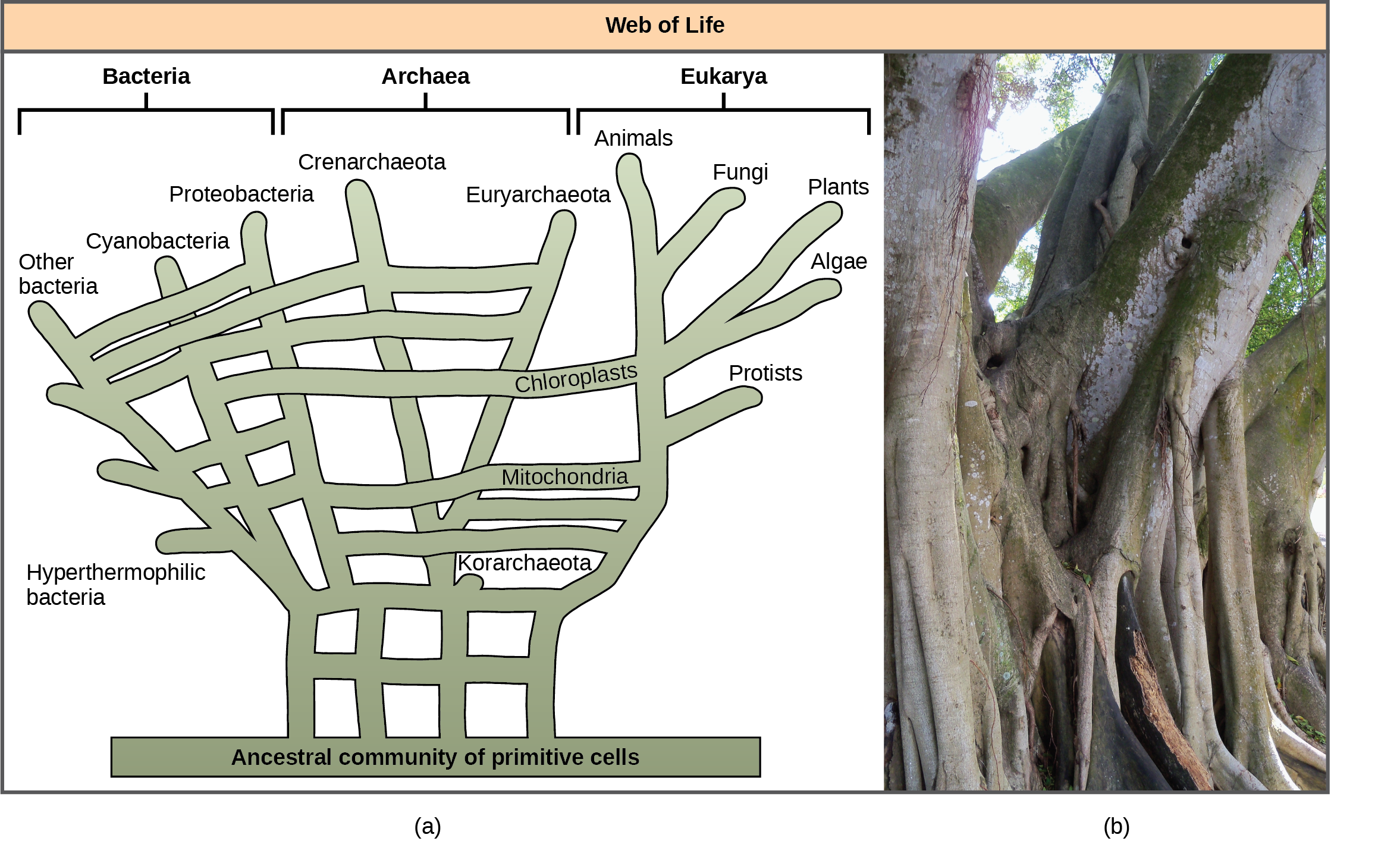
Recognizing the importance of HGT, especially in prokaryote evolution, has caused some to propose abandoning the classic “tree of life” model. In 1999, W. Ford Doolittle proposed a phylogenetic model that resembles a web or a network more than a tree. The hypothesis is that eukaryotes evolved not from a single prokaryotic ancestor, but from a pool of many species that were sharing genes by HGT mechanisms. As Figure 5a shows, some individual prokaryotes were responsible for transferring the bacteria that caused mitochondrial development to the new eukaryotes; whereas, other species transferred the bacteria that gave rise to chloroplasts. Scientists often call this model the “web of life.” In an effort to save the tree analogy, some have proposed using the Ficus tree (Figure 5b) with its multiple trunks as a phylogenetic way to represent a diminished evolutionary role for HGT.
Ring of Life Models
Others have proposed abandoning any tree-like model of phylogeny in favor of a ring structure, the so-called “ring of life” (Figure 6). This is a phylogenetic model where all three domains of life evolved from a pool of primitive prokaryotes. Lake, again using the conditioned reconstruction algorithm, proposes a ring-like model in which species of all three domains—Archaea, Bacteria, and Eukarya—evolved from a single pool of gene-swapping prokaryotes. His laboratory proposes that this structure is the best fit for data from extensive DNA analyses performed in his laboratory, and that the ring model is the only one that adequately takes HGT and genomic fusion into account. However, other phylogeneticists remain highly skeptical of this model.
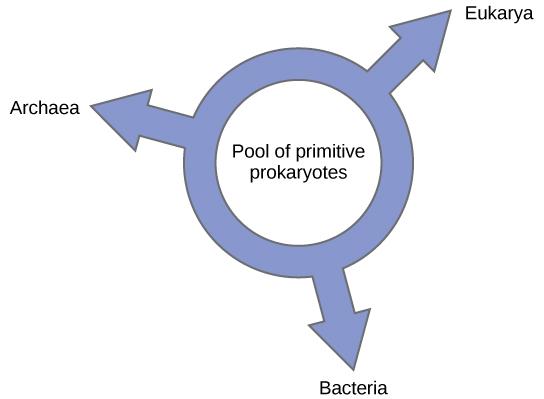
In summary, we must modify Darwin’s “tree of life” model to include HGT. Does this mean abandoning the tree model completely? Even Lake argues that scientists should attempt to modify the tree model to allow it to accurately fit his data, and only the inability to do so will sway people toward his ring proposal.
This doesn’t mean a tree, web, or a ring will correlate completely to an accurate description of phylogenetic relationships of life. A consequence of the new thinking about phylogenetic models is the idea that Darwin’s original phylogenetic tree concept is too simple, but made sense based on what scientists knew at the time. However, the search for a more useful model moves on: each model serves as hypotheses to test with the possibility of developing new models. This is how science advances. Researchers use these models as visualizations to help construct hypothetical evolutionary relationships and understand the massive amount of data that requires analysis.
Glossary
- eukaryote-first hypothesis
- proposal that prokaryotes evolved from eukaryotes
- gene transfer agent (GTA)
- bacteriophage-like particle that transfers random genomic segments from one species of prokaryote to another
- genome fusion
- fusion of two prokaryotic genomes, presumably by endosymbiosis
- horizontal gene transfer (HGT)
- (also, lateral gene transfer) transfer of genes between unrelated species
- mitochondria-first hypothesis
- proposal that prokaryotes acquired a mitochondrion first, followed by nuclear development
- nucleus-first hypothesis
- proposal that prokaryotes acquired a nucleus first, and then the mitochondrion
- ring of life
- phylogenetic model where all three domains of life evolved from a pool of primitive prokaryotes
- web of life
- phylogenetic model that attempts to incorporate the effects of horizontal gene transfer on evolution
Access for free at https://openstax.org/books/biology-2e/pages/1-introduction
