64 Structure of Prokaryotes: Bacteria and Archaea
By the end of this section, you will be able to do the following:
- Describe the basic structure of a typical prokaryote
- Describe important differences in structure between Archaea and Bacteria
There are many differences between prokaryotic and eukaryotic cells. The name “prokaryote” suggests that prokaryotes are defined by exclusion—they are not eukaryotes, or organisms whose cells contain a nucleus and other internal membrane-bound organelles. However, all cells have four common structures: the plasma membrane, which functions as a barrier for the cell and separates the cell from its environment; the cytoplasm, a complex solution of organic molecules and salts inside the cell; a double-stranded DNA genome, the informational archive of the cell; and ribosomes, where protein synthesis takes place. Prokaryotes come in various shapes, but many fall into three categories: cocci (spherical), bacilli (rod-shaped), and spirilli (spiral-shaped) (Figure 1).

The Prokaryotic Cell
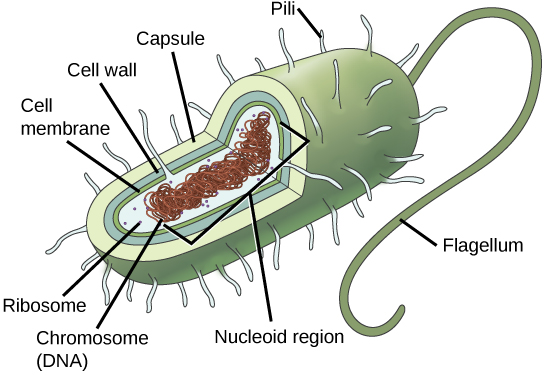
Recall that prokaryotes are unicellular organisms that lack membrane-bound organelles or other internal membrane-bound structures (Figure 2). Their chromosome—usually single—consists of a piece of circular, double-stranded DNA located in an area of the cell called the nucleoid. Most prokaryotes have a cell wall outside the plasma membrane. The cell wall functions as a protective layer, and it is responsible for the organism’s shape. Some bacterial species have a capsule outside the cell wall. The capsule enables the organism to attach to surfaces, protects it from dehydration and attack by phagocytic cells, and makes pathogens more resistant to our immune responses. Some species also have flagella (singular, flagellum) used for locomotion, and pili (singular, pilus) used for attachment to surfaces including the surfaces of other cells. Plasmids, which consist of extra-chromosomal DNA, are also present in many species of bacteria and archaea.
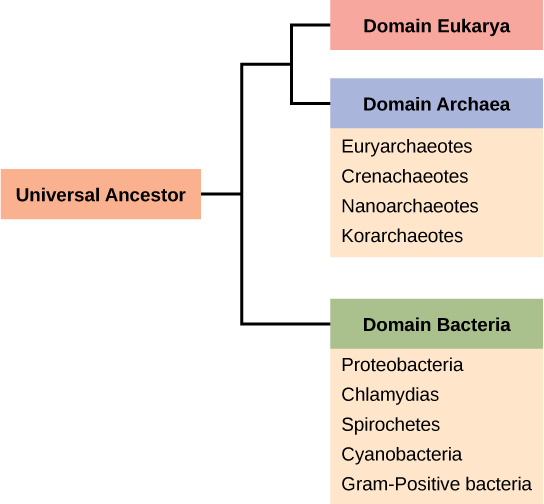
Recall that prokaryotes are divided into two different domains, Bacteria and Archaea, which together with Eukarya, comprise the three domains of life (Figure 3).
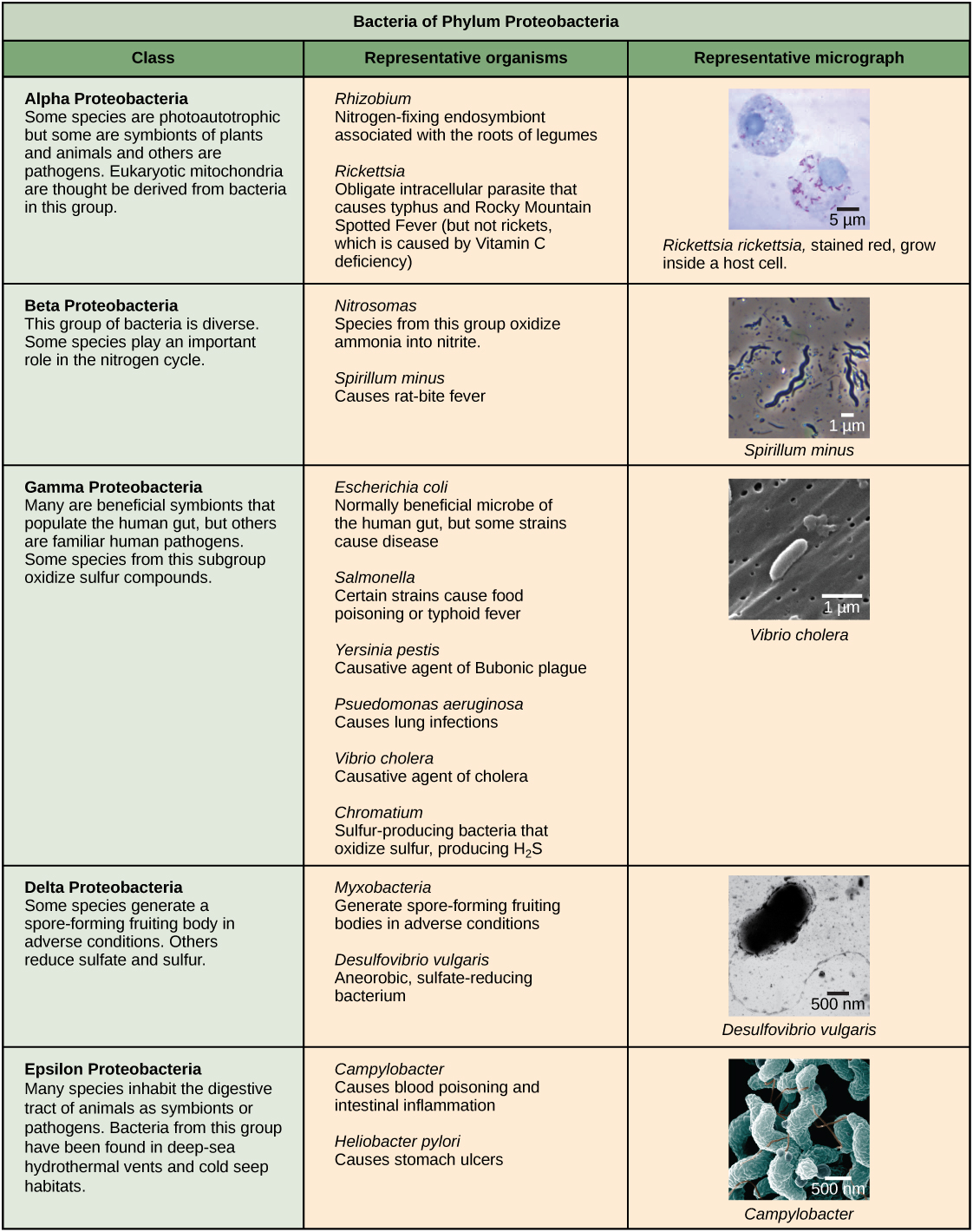
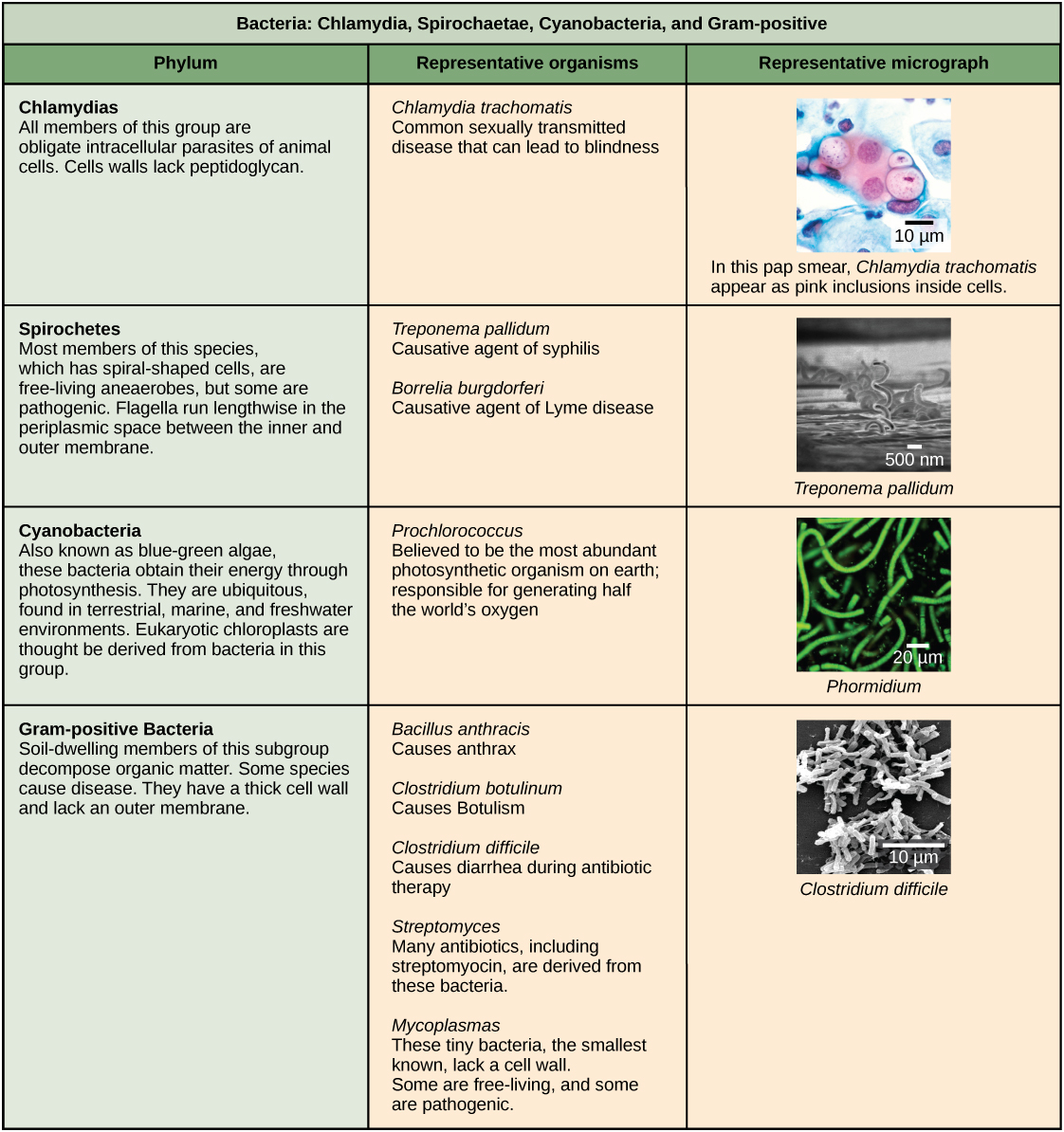
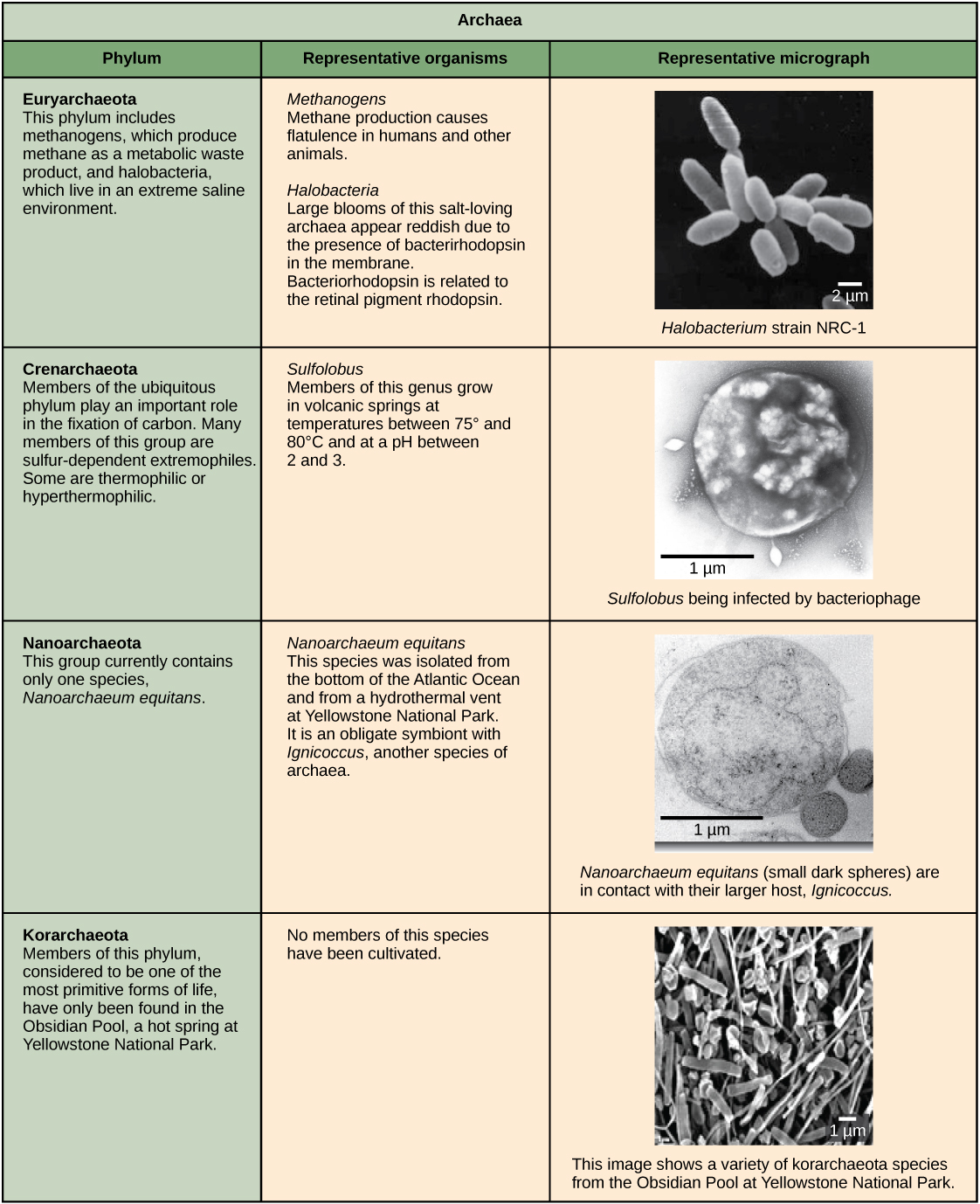
Characteristics of bacterial phyla are described in Figure 4 and Figure 5. Major bacterial phyla include the Proteobacteria, the Chlamydias, the Spirochaetes, the photosynthetic Cyanobacteria, and the Gram-positive bacteria. The Proteobacteria are in turn subdivided into several classes, from the Alpha- to the Epsilon proteobacteria. Eukaryotic mitochondria are thought to be the descendants of alphaproteobacteria, while eukaryotic chloroplasts are derived from cyanobacteria. Archaeal phyla are described in Figure 6.
The Plasma Membrane of Prokaryotes
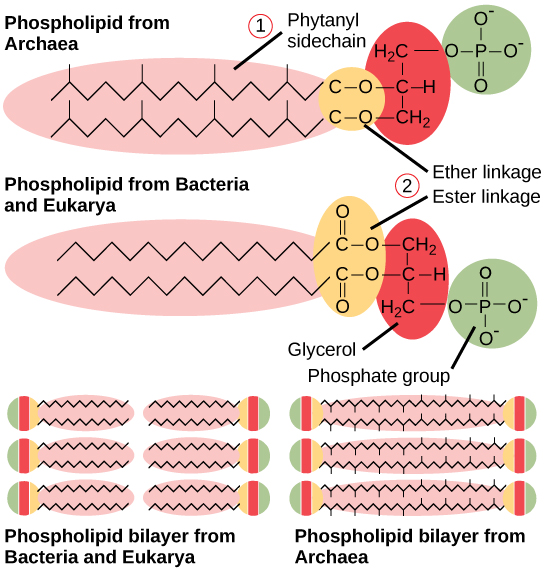
The prokaryotic plasma membrane is a thin lipid bilayer (6 to 8 nanometers) that completely surrounds the cell and separates the inside from the outside. Its selectively permeable nature keeps ions, proteins, and other molecules within the cell and prevents them from diffusing into the extracellular environment, while other molecules may move through the membrane. Recall that the general structure of a cell membrane is a phospholipid bilayer composed of two layers of lipid molecules. In archaeal cell membranes, isoprene (phytanyl) chains linked to glycerol replace the fatty acids linked to glycerol in bacterial membranes. Some archaeal membranes are lipid monolayers instead of bilayers (Figure 7).
The Cell Wall of Prokaryotes
The cytoplasm of prokaryotic cells has a high concentration of dissolved solutes. Therefore, the osmotic pressure within the cell is relatively high. The cell wall is a protective layer that surrounds some cells and gives them shape and rigidity. It is located outside the cell membrane and prevents osmotic lysis (bursting due to increasing volume). The chemical composition of the cell wall varies between Archaea and Bacteria, and also varies between bacterial species.
Bacterial cell walls contain peptidoglycan, composed of polysaccharide chains that are cross-linked by unusual peptides containing both L- and D-amino acids including D-glutamic acid and D-alanine. (Proteins normally have only L-amino acids; as a consequence, many of our antibiotics work by mimicking D-amino acids and therefore have specific effects on bacterial cell-wall development.) There are more than 100 different forms of peptidoglycan. S-layer (surface layer) proteins are also present on the outside of cell walls of both Archaea and Bacteria.
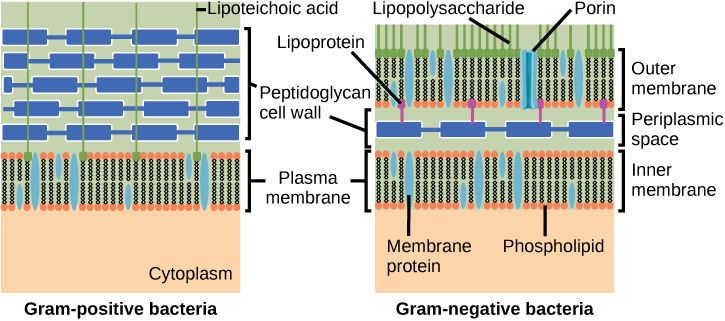
Bacteria are divided into two major groups: Gram positive and Gram negative, based on their reaction to Gram staining. Note that all Gram-positive bacteria belong to one phylum; bacteria in the other phyla (Proteobacteria, Chlamydias, Spirochetes, Cyanobacteria, and others) are Gram-negative. The Gram staining method is named after its inventor, Danish scientist Hans Christian Gram (1853–1938). The different bacterial responses to the staining procedure are ultimately due to cell wall structure. Gram-positive organisms typically lack the outer membrane found in Gram-negative organisms (Figure 8). Up to 90 percent of the cell-wall in Gram-positive bacteria is composed of peptidoglycan, and most of the rest is composed of acidic substances called teichoic acids. Teichoic acids may be covalently linked to lipids in the plasma membrane to form lipoteichoic acids. Lipoteichoic acids anchor the cell wall to the cell membrane. Gram-negative bacteria have a relatively thin cell wall composed of a few layers of peptidoglycan (only 10 percent of the total cell wall), surrounded by an outer envelope containing lipopolysaccharides (LPS) and lipoproteins. This outer envelope is sometimes referred to as a second lipid bilayer. The chemistry of this outer envelope is very different, however, from that of the typical lipid bilayer that forms plasma membranes.
VISUAL CONNECTION
Which of the following statements is true?
- Gram-positive bacteria have a single cell wall anchored to the cell membrane by lipoteichoic acid.
- Porins allow entry of substances into both Gram-positive and Gram-negative bacteria.
- The cell wall of Gram-negative bacteria is thick, and the cell wall of Gram-positive bacteria is thin.
- Gram-negative bacteria have a cell wall made of peptidoglycan, whereas Gram-positive bacteria have a cell wall made of lipoteichoic acid.
Answer:
Statement “a” is true.
Archaean cell walls do not have peptidoglycan. There are four different types of archaean cell walls. One type is composed of pseudopeptidoglycan, which is similar to peptidoglycan in morphology but contains different sugars in the polysaccharide chain. The other three types of cell walls are composed of polysaccharides, glycoproteins, or pure protein. Other differences between Bacteria and Archaea are seen in Table 1. Note that features related to DNA replication, transcription and translation in Archaea are similar to those seen in eukaryotes.
| Structural Characteristic | Bacteria | Archaea |
|---|---|---|
| Cell type | Prokaryotic | Prokaryotic |
| Cell morphology | Variable | Variable |
| Cell wall | Contains peptidoglycan | Does not contain peptidoglycan |
| Cell membrane type | Lipid bilayer | Lipid bilayer or lipid monolayer |
| Plasma membrane lipids | Fatty acids-glycerol ester | Phytanyl-glycerol ethers |
| Chromosome | Typically circular | Typically circular |
| Replication origins | Single | Multiple |
| RNA polymerase | Single | Multiple |
| Initiator tRNA | Formyl-methionine | Methionine |
| Streptomycin inhibition | Sensitive | Resistant |
| Calvin cycle | Yes | No |
Reproduction
Reproduction in prokaryotes is asexual and usually takes place by binary fission. (Recall that the DNA of a prokaryote is a single, circular chromosome.) Prokaryotes do not undergo mitosis; instead, the chromosome is replicated and the two resulting copies separate from one another, due to the growth of the cell. The prokaryote, now enlarged, is pinched inward at its equator and the two resulting cells, which are clones, separate. Binary fission does not provide an opportunity for genetic recombination or genetic diversity, but prokaryotes can share genes by three other mechanisms.
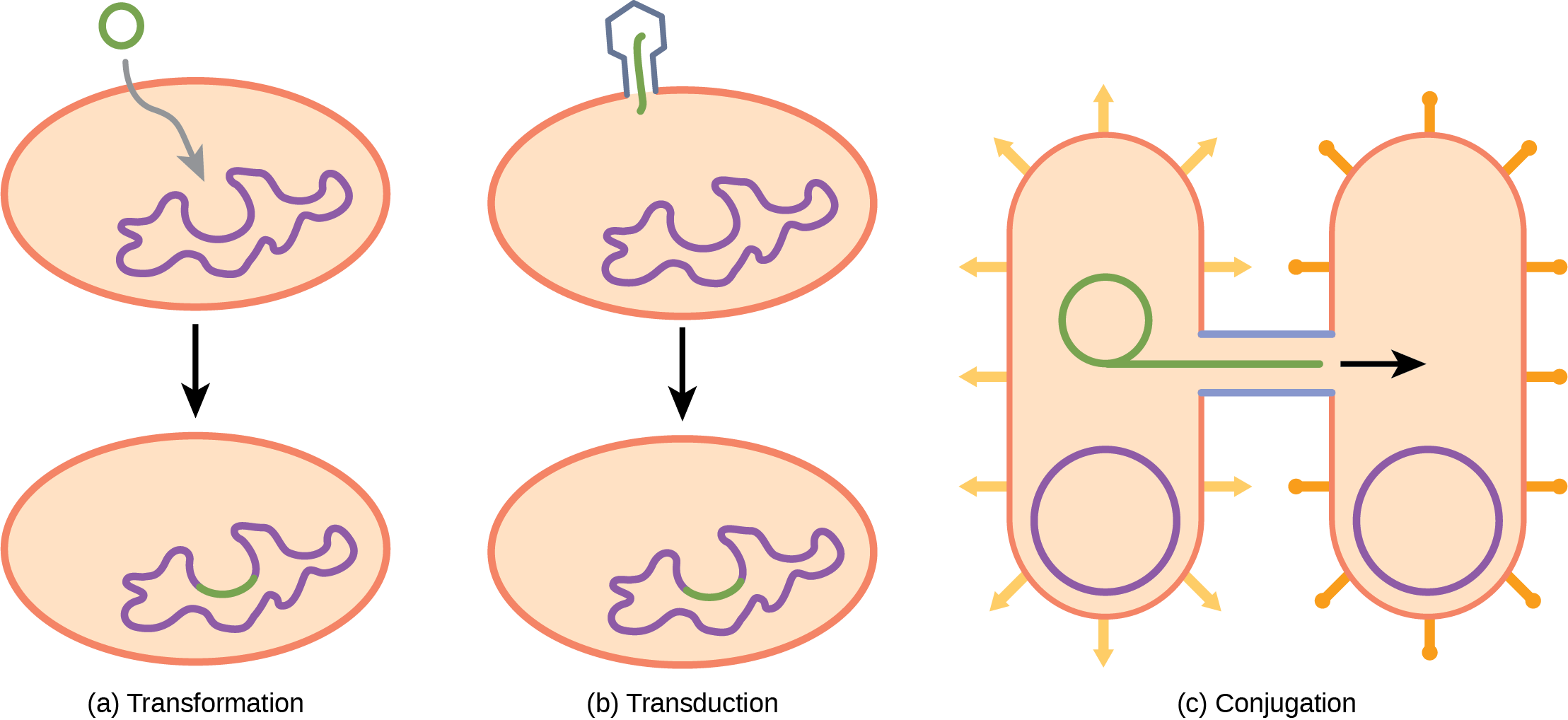
In transformation, the prokaryote takes in DNA shed by other prokaryotes into its environment. If a nonpathogenic bacterium takes up DNA for a toxin gene from a pathogen and incorporates the new DNA into its own chromosome, it too may become pathogenic. In transduction, bacteriophages, the viruses that infect bacteria, may move short pieces of chromosomal DNA from one bacterium to another. Transduction results in a recombinant organism. Archaea also have viruses that may translocate genetic material from one individual to another. In conjugation, DNA is transferred from one prokaryote to another by means of a pilus, which brings the organisms into contact with one another, and provides a channel for transfer of DNA. The DNA transferred can be in the form of a plasmid or as a composite molecule, containing both plasmid and chromosomal DNA. These three processes of DNA exchange are shown in Figure 9.
Reproduction can be very rapid: a few minutes for some species. This short generation time coupled with mechanisms of genetic recombination and high rates of mutation result in the rapid evolution of prokaryotes, allowing them to respond to environmental changes (such as the introduction of an antibiotic) very quickly.
EVOLUTION CONNECTION
The Evolution of Prokaryotes
How do scientists answer questions about the evolution of prokaryotes? Unlike with animals, artifacts in the fossil record of prokaryotes offer very little information. Fossils of ancient prokaryotes look like tiny bubbles in rock. Some scientists turn to genetics and to the principle of the molecular clock, which holds that the more recently two species have diverged, the more similar their genes (and thus proteins) will be. Conversely, species that diverged long ago will have more genes that are dissimilar.
Scientists at the NASA Astrobiology Institute and at the European Molecular Biology Laboratory collaborated to analyze the molecular evolution of 32 specific proteins common to 72 species of prokaryotes.1 The model they derived from their data indicates that three important groups of bacteria—Actinobacteria, Deinococcus, and Cyanobacteria (collectively called Terrabacteria by the authors)—were the first to colonize land. Actinobacteria are a group of very common Gram-positive bacteria that produce branched structures like fungal mycelia, and include species important in decomposition of organic wastes. You will recall that Deinococcus is a genus of bacterium that is highly resistant to ionizing radiation. It has a thick peptidoglycan layer in addition to a second external membrane, so it has features of both Gram-positive and Gram-negative bacteria.
Cyanobacteria are photosynthesizers, and were probably responsible for the production of oxygen on the ancient earth. The timelines of divergence suggest that bacteria (members of the domain Bacteria) diverged from common ancestral species between 2.5 and 3.2 billion years ago, whereas the Archaea diverged earlier: between 3.1 and 4.1 billion years ago. Eukarya later diverged from the archaean line. The work further suggests that stromatolites that formed prior to the advent of cyanobacteria (about 2.6 billion years ago) photosynthesized in an anoxic environment and that because of the modifications of the Terrabacteria for land (resistance to drying and the possession of compounds that protect the organism from excess light), photosynthesis using oxygen may be closely linked to adaptations to survive on land.
Footnotes
- 1 Battistuzzi, FU, Feijao, A, and Hedges, SB. A genomic timescale of prokaryote evolution: Insights into the origin of methanogenesis, phototrophy, and the colonization of land. BioMed Central: Evolutionary Biology 4 (2004): 44, doi:10.1186/1471-2148-4-44.
- capsule
- external structure that enables a prokaryote to attach to surfaces and protects it from dehydration
- conjugation
- process by which prokaryotes move DNA from one individual to another using a pilus
- Gram negative
- bacterium whose cell wall contains little peptidoglycan but has an outer membrane
- Gram positive
- bacterium that contains mainly peptidoglycan in its cell walls
- peptidoglycan
- material composed of polysaccharide chains cross-linked to unusual peptides
- pilus
- surface appendage of some prokaryotes used for attachment to surfaces including other prokaryotes
- pseudopeptidoglycan
- component of archaea cell walls that is similar to peptidoglycan in morphology but contains different sugars
- S-layer
- surface-layer protein present on the outside of cell walls of archaea and bacteria
- teichoic acid
- polymer associated with the cell wall of Gram-positive bacteria
- transduction
- process by which a bacteriophage moves DNA from one prokaryote to another
- transformation
- process by which a prokaryote takes in DNA found in its environment that is shed by other prokaryotes
Glossary
Access for free at https://openstax.org/books/biology-2e/pages/1-introduction